Користувач:TnoXX/Чернетка
Алярна та базальна пластинки, які простягаються уздовж ЦНС, та невромери, які ділять нервову систему у дорсально-вентральному напрямку, разом визначають розвиток ядер черепних нервів:
| Невромер | Похідне базальної пластинки | Похідне алярної пастинки | Примітки |
|---|---|---|---|
| Мезомер 1 | Ядро окорухового нерва, додаткове ядро окорухового нерва | — | |
| Невромер перешийка | Ядро блокового нерва | — | [1] |
| Ромбомер 2 | Рухове ядро трійчастого нерва | — | |
| Ромбомер 3 | — | Головне ядро трійчастого нерва | |
| Ромбомер 4 | Спинномозкове ядро трійчастого нерва | ||
| Ромбомер 5 | Спинномозкове ядро трійчастого нерва | ||
| Ромбомер 6 | Спинномозкове ядро трійчастого нерва | ||
| Ромбомер 7 | Подвійне ядро | Спинномозкове ядро трійчастого нерва | |
| Ромбомер 8 | Подвійне ядро | Спинномозкове ядро трійчастого нерва | |
| Ромбомер 9 | Подвійне ядро, ядро під'язикового нерва | Спинномозкове ядро трійчастого нерва | |
| Ромбомер 10 | ядро під'язикового нерва | Спинномозкове ядро трійчастого нерва | |
| Ромбомер 11 | ядро під'язикового нерва | Спинномозкове ядро трійчастого нерва |
Регенерація зорового нерва[ред. | ред. код]
Атрофія зорового нерва веде до втрати зору. Зважаючи на важливість зору для людини, питання регенерації зорового нерва має високу клінічну важливість. Регенерація зорового нерва ускладнена тим, що, будучи гістологічно складовою ЦНС, він має обмежені можливості до відновлення у порівнянні із ПНС.
Для людини
Неоліт та давній світ[ред. | ред. код]
Мікроанатомія нервових утворів[ред. | ред. код]
Біла та сіра речовини[ред. | ред. код]

Біла та сіра речовини так названі через свій колір, який можна спостерігати при розрізі утворів ЦНС, наприклад, головного мозку. Біла речовина складається з мієлінізованих аксонів, які в ЦНС утворюють нервові шляхи. Сіра речовина представлена перикаріонами та дендритами нейронів і утворює нервові центри, які бувають двох типів: центри екранного типу представляють собою кору кінцевого мозку та мозочка; центри ядерного типу – це всі ядра ЦНС, які являють собою компакто розташовані ділянки сірої речовини серед білої речовини.[2][3]
Нерви[ред. | ред. код]

Нерв (лат. nervus) є периферичною нервовою структурою, основою якої є мієлінізовані або немієлінізовані аксони. У випадку мієлінізованого аксона до складу нерва також входять шваннівські клітини; периневрій та ендоневрій також мають свій клітинний апарат. Кожен нерв має судини, які необхідні для його живлення. Кожен аксон (в контексті нерва його можна називати нервовим волокном) покритий сполучнотканниною оболонкою – ендоневрієм. Декілька нервових волокон оточені іншою сполучнотканниною оболонкою – периневрієм – який формує з нервових волокон нервові пучки. Окрім формування пучків, периневрій створює периневральний бар'єр, який обмежує доступ до нервових волокон і служить для підтримки гомеостазу в нерві. Нервові пучки оточені епіневрієм – зовнішньою сполучнотканниною оболонкою, яка власне формує нерв. Кількість пучків визначає товщину нерва.[4]
Нерви можуть бути чутливими (вісцеро- та соматочутливими), руховими (вісцеро- та соматомоторними), а також змішаними (наявні як рухові, так і чутливі волокна). За натомічним принципом вони бувають черепними та спинномозковими.
Нервовий вузол[ред. | ред. код]

Нервовий вузол (лат. ganglion) – це скупчення нейронів, мантійних гліоцитів та їх відростків, які є складовою ПНС. Нервові вузли є аналогами ядер на периферії. Вони можуть бути чутливими (спинномозкові та ті, що належать черепним нервам) та вегетативними (симптичними та парасимпатичними).
Кожен чутливий вузол вкритий сполучнотканниною оболонкою, від якої в середину відходять сполучнотканинні перегородки. У всіх чутливих вузлах містяться тільки псевдоуніполярні нейрони (виняток – вузли VIII пари черепних нервів). Тіла нейронів розташовані периферично по відношенню до їх відростків, які містяться в центрі вузла.
Вегетативні вузли також вкриті сполучнотканинною оболонкою. Вони мають дві відмінності, якщо порівнювати з чутливими вузлами: нейрони цих вузлів є мультиполярними, а волокна не займають центральне положення, як у чутливих вузлах.
Глія[ред. | ред. код]
Нейроглія ЦНС[ред. | ред. код]
До нейроглії ЦНС відносять астроглію, яка є найбільш неоднорідною глією в ЦНС, NG-2-глію, олігодендритоглію та мікроглію[5].
Астроглія має ряд важливих функцій, серед яких забезпечення адекватної кількості йонів, води та інших речовин в позаклітинному середовищі нейронів, контроль за мікроциркуляцією, синхронізація роботи нейронів, видалення нейротрансмітерів зі синаптичної щілини, регуляція синаптогенезу, утворення гемато-енцефалічного бар'єру, забезпечення міграції нейронів під час внутрішньоутробного розвитку нервової системи та забезпечення правильної цитоархітектури сірої речовини, вистиляє шлуночки та контактує зі спинномозковою рідиною[6][7][8].
Сама астроглія складається з великої кількості морфологічно відмінних клітин та їх підтипів. До астроглії відносять різні астроцити (фіброзні, протоплазматичні, крайові), клітини Бергмана та клітини Мюллера, клітини судинних сплетень, епендимоцити, таніцити, пітуїцити, пігментні клітини сітківки[9].
Олігодендритоглія складається з мієлінізуючих олігодендроцитів[10] та двох підтипів немієлінізуючих олігодендроцитів: периневрональних олігодендроцитів, функція яких залишається невідомою, та поліденроцитів, які згідно сучасних поглядів, відносяться до окремого четвертого класу глії ЦНС – NG-2-глії[11]. Мієлінізуючі олігодедроцити мають велику кількість відростків, які оточують багато аксонів та продукують мієлін. Окрім того, один аксон вони можуть оточувати в багатьох місцях[12][13].
NG-2-глія, або полідендритоглія, є наймолодшим окремо виділеним класом нейроглії. Полідендритоцити за своєю суттю є попередниками олігодендритоцитів і здатні перетворюватися в них протягом всього життя людини. Проте лише незначна частина цих клітин генерує нові олігодендроцити. Окрім того ці клітини здатні утворювати синапси зі нейронами, що є унікальним випадком для нейроглії (тому їхня ще одна назва – синапсоцити).[14]
Мікроглія за своїм походженням є відмінною від усієї решти нейроглії. Моноцити, які проникають у паренхіму головного мозку протягом ранніх стадій його розвитку, складають подальшу клітинну основу мікроглії[15]. Основна функція мікроглії – імунна: фагоцитуванні клітинного «сміття», яке виникає при ушкодженні ЦНС та презентація антигенів імунокомпетентним клітинам. Окрім того мікроглія відіграє важливу роль в дозріванні нервової системи, утворенні нервових шляхів, ангіогенезі ЦНС.[16][17]
Нейроглія ПНС[ред. | ред. код]
До периферичної нейроглії відносять шванівські клітини, мантійні гліоцити, які розташовані в нервових вузлах, ентеричні гліальні клітини та нюхові оболонкові клітини.
Шванівські клітини, або лемоцити, бувають трьох видів: мієлінізуючі, немієлінізуючі та перисинаптичні. Перші охоплюють аксон та продукують мієлін (на відміну від олігодендроцита, вони охоплюють лише один сегмент аксона)[18]. Немієлінізуючі шванівські клітини оточують немієлінізовані аксони, при цьому мієлін вони не виробляють. Перисинаптичні лемоцити покривають терміналі аксона.[19]
Функція мантійних гліоцитів до кінця не вивчена, однак вважається, що вони виконують функцію, подібну до астроцитів в ЦНС, тобто підтримують оптимальні умови для функціонування нейронів у нервових вузлах[20].
Ентерична глія забезпечує підтримання гомеостазу в ентеричній нервовій системі, на ряду з імунними клітинами та епітеліоцитами слизової оболонки виконує бар'єрні функції[21].
Оболонкові нюхові клітини виконують виконують подібні функції до двох попередніх глій (ізоляція та контроль за гомеостазом), а також забезпечують можливість регенерації нейронів в цій ділянці[22].
Аксонний транспорт[ред. | ред. код]
Транспорт речовин або органел вздовж аксона називається аксонним транспортом[23].
Якщо речовина транспортується на периферію, то такий транспорт називається антеградним. Антеградний транспорт буває швидким та повільним.[24] Завдяки антеградному швидкому транспорту речовини проходять відстань приблизно у 100–400 мм[25] на день. До таких речовин можна віднести мітохондрії, білки синаптичних міхурців, мембранні білки.[23] Найважливішою складовою цитоскелету, яка забезпечує швидкий антеградний транспорт, є кінезин-1 з родини кінезинів[26][27]. Повільний транспорт відбувається зі швидкістю 0,2-6 мм[23] на день. Завдяки ньому транспортуються елементи цитоскелету та розчинені білки[23][28]. Цитоплазматичний динеїн є найважливішою складовою цитоскелета у забезпеченні повільного транспорта[26].
Частина синтезованих сполук повертається по аксону до перикаріону. Це — ретроградний аксонний транспорт, який має тільки швидку складову. Такий транспорт необхідний для утилізації та деградації непотрібних речовин, для ресинтезу необхідних сполук та захоплення нейротрофічних факторів — речовин, необхідних для нормального функціонування нейрона.[23][24][29] Завдяки такому транспорту деякі віруси та токсини здатні проникати в нейрони[23]. Головну роль в забезпеченні такого транспорту відіграє той же цитоплазматичний динеїн[26][27].
Наявність цілого аксона, по якому може відбуватися аксонний транспорт, є умовою функціонування аксона; у випадку перерізання аксона його дистальна частина піддається дегенерації (волерівська дегенерація). З іншого боку цілий аксон необхідний для існування нейрона, адже, як вже було сказано, нейротрофічні фактори прямують до перикаріона з периферії.[29]
Макроанатомія[ред. | ред. код]
У людини нервову систему за анатомічним принципом ділять на центральну (центральна нервова система, або ЦНС) та периферичну (периферична нервова система, або ПНС). Центральна нервова система, або невраксіс, складається з двох великих частин – спинного мозку (лат. medulla spinalis) та головного мозку (лат. encephalon). Периферична – з черепних та спинномозкових нервів (які також можуть утворювати різні сплетення) та нервових вузлів.[30]
Спинний мозок[ред. | ред. код]

Спинний мозок являє собою валик, який у дорослої людини простягається від перехрестя пірамід (лат. decussatio pyramidum) та місця виходу I пари спинномозкових нервів до рівня другого поперекового хребця[31]. Спинний мозок поділений на відповідні сегменти (8 шийних, 12 грудних, 5 поперекових, 5 крижових та 1-3 куприкових сегменти)[32]. Товщина спинного мозку дорівнює приблизно 1-1,5 см, однак наявні два потовщення (шийне (лат. intumescentia cervicalis) та попереково-крижове (лат. intumescentia lumbosacralis)). Спинний мозок має порожнину – центральний канал (лат. canalis centralis), який є частиною шлуночкової системи і в якому циркулює спинномозкова рідина. Довкола центрального каналу розміщена сіра речовина спинного мозку (лат. substantia grisea), яка на поперечному зрізі має вигляд метелика.[33] В цьому метелику розрізняють передній (лат. cornu anterius) та задній (лат. cornu posterius) роги, а також бічний ріг (лат. cornu laterale), який трапляється тільки від C8 до L3 сегментів спинного мозку.[34] Якщо брати зріз в сагітальній площині, то по обидва боки цетрального каналу розміщені дві колони сірої речовини, об'єднані невеликою пластинкою сірої речовини безпосередньо у ділянці центрального каналу. Кожна з колон поділена на три стовпи: передній (лат. columna anterior), який відповідає передньому рогу, проміжний (лат. columna intermedia), який простягається від C8 до L3 та відповідає бічному рогу, та задній (лат. columna posterior), який відповідає однойменному рогу.
В передніх рогах в основному лежать соматомотонейрони (α- та γ-мотонейони)[35], хоча у крижових сегментах (S2 – S4) тут також розміщені парасимпатичні ядра, які складаються з вісцеромотонейронів; у бічних рогах – розміщені симпатичні ядра та ядра вісцеральної чутливості; у задніх рогах – розміщені чутливі ядра[36].
Біла речовина спинного мозку розміщена поверхнево відносно сірої. Борознами вона поділена на три канатики – передній (лат. funiculus anterior), бічний (лат. funiculus lateralis) та задній (лат. funiculus posterior)[37]. В кожному з канатиків проходять нервові шляхи. Так, основну частину заднього кантика складають тонкий (лат. fasciculus gracilis) та клиноподібний пучки (лат. fasciculus cuneatus), які пов'язані з пропріоцептивною чутливістю[38]. В середньому канатику міститься бічний кірково-спинномозговий шлях (лат. tractus corticospinalis lateralis), бічний спинномозково-таламічний шлях (лат. tractus spinothalamicus lateralis), ряд екстрапірамідних шляхів та шляхів власного сегментарного апарату спинного мозку[39]. У передньому канатику міститься передній спинномозково-таламічний шлях (лат. tractus spinothalamicus anterior), передній кірково-спинномозковий шлях (лат. tractus corticospinalis anterior) та ряд екстрапірамідних шляхів[40].
Головний мозок[ред. | ред. код]

Головний мозок є тою частиною ЦНС, яка розміщена в порожнині черепа[41]. Він складається з шести великих відділів: довгастого мозку (лат. medulla oblongata), моста (лат. pons), середнього мозку (лат. mesencephalon), проміжного мозку (лат. diencephalon), кінцевого мозку (лат. telencephalon) та мозочка (лат. cerebellum). Перші три відділи утворюють стовбур мозку (лат. truncus cerebri).[42] Кожний з цих віддів має порожнину, яка входить до шлуночкової системи головного мозку.
Довгастий мозок складається з висхідних та нисхідних шляхів, а також з розміщений серед цієї білої речовини «острівців» сірої речовини – ядер черепних нервів (IX-XII) та деяких інших ядер[43]. В довгастому мозку розміщені центри дихання, ковтання, блювання та серцево-судинний центр. Іншою сірою речовиною є сітчасте утворення (лат. formatio reticularis), яке простягається по всьому стовбуру мозку[44].
Міст також складається з великої кількості провідних шляхів, містить ядра черепних нервів (V-VIII) та сітчасте утворення[45]. Разом з довгастим мозком утворюють задній мозок (лат. metencephalon), який має порожнину – четвертий шлуночок (лат. ventriculus quartus).[46]
Мозочок є тою частиною головного мозку, яку пов'язують з руховою сферою, а останнім часом – з процесами пам'яті, уваги, мовлення, планування. Складається з тіла мозочка (лат. corpus cerebelli), розділеного на черв'як мозочка (лат. vermis cerebelli) та дві півкулі мозочка (лат. hemisperia cerebelli), та клаптиково-вузлову частку (лат. lobus flocullonodularis).[47]
Середній мозок окрім провідних шляхів містить ядра III-IV пар черепних нервів, підкіркові центри зору та слуху, важливі екстрапірамідні ділянки (чорну речовину (лат. substantia nigra) та червоне ядро (лат. nucleus ruber)). Також має порожнину – водопровід середнього мозку (лат. aqueductus mesencephali), який сполучає IV та III шлуночки.[48]
Проміжний мозок має складну будову і згідно з сучасною анатомічною номенклатурою складається з таламуса, епіталамуса, гіпоталамуса, субталамуса та метаталамуса.[49]. Порожниною проміжного мозку є третій шлуночок (лат. ventriculus tertius)[50].
Кінцевий мозок складається з двох півкуль (лат. hemispheria cerebri), об'єднаних мозолистим тілом (лат. corpus callosum). Поверхня півкуль покрита корою (лат. cortex). В глибині півкуль розміщена біла речовина (уся сукупність шляхів, що прямують до півкуль) та базальні ядра (лат. nuclei basales). Кожна півкуля має порожнину – бічний шлуночок (лат. ventriculus lateralis).[51]
Периферична нервова система[ред. | ред. код]
Усі ті нервові утвори, які розміщені поза ЦНС відносять до периферичної нервової системи. У людини наявні 12 пар черепних нервів (разом з термінальним нервом, їх є 13 пар)[52] та 31-33 (залежно від кількості сегментів спинного мозку) пар спинномозкових нервів[32]. З нервами також пов'язані нервові вузли і сплетення[53].
Спинномозковий нерв (лат. nervus spinalis) утворений двома корінцями, які виходять зі спинного мозку і пізніше утворюють єдиний стовбур: переднього рухового корінця (лат. radix anterior (motoria)) та заднього чутливого (лат. radix posterior (sensoria)). Передній корінець у різних сегментах спинного мозку може складатися з соматомоторних та вісцеромоторних волокон. Задній корінець пов'язаний з чутливим вузлом (лат. ganglion sensorium nervi spinalis), який цей корінець в основному і утворює, та може складатися з волокон соматичної та вісцеральної чутливості (залежно від присутності певних ядер в сегменті спинного мозку)[32]. Спинномозкові нерви утворюють великі сплетення – шийне (лат. plexus cervicalis)[54], плечове (лат. plexus brachialis)[55], поперекове (лат. plexus lumbalis)[56], крижове (лат. plexus sacralis)[57] та куприкове (лат. plexus coccygeus)[58].
Окрім чутливих вузлів виділяють симпатичні нервові вузли: прихребтові, які є складовою парного симпатичного стовбура (лат. truncus sympathicus), аксони яких приєднюються до спинномозкового нерва та прямують на периферію з метою іннервації органа-мішені[59] та передхребтові (лат. ganglia prevertebrales), які входять до складу нутрощевих сплетень та є їх симпатичною складовою[60].
Нутрощеві сплетення у людини складаються з нутрощевих вузлів (передхребтових, позаорганних та внутрішньорганних), перед- та завузлових симпатичних та парасиматичних волокон і представлені[61]:
- черепно-шийною частиною (загальне сонне сплетення, внутрішнє сонне сплетення, печеристе сплетення, зовнішнє соннє сплетення, автономне плечове сплетення, хребтове сплетення)
- грудної частини (грудне аортальне сплетення, серцеве сплетення, стравохідне сплетення, легеневе сплетення)
- черевною частиною, у якої є велика кількість сплетень, серед яких найважливішими виступають черевне аортальне та черевне (сонячне) сплетення
- тазовою частиною (верхнє підчеревне сплетення та нижнє підчеревне сплетення)
Окремо, насаперед з точки зору фізіології, виділяють сплетення, які є складовою так званої метасимпатичної нервової системи. Такі сплетення хоч і пов'язані з ЦНС, однак мають власний рефлекторний апарат і володіють великою функціональною автономією. Найбільш вивченою складовою цієї системи є ентерична нервова система, представлена сплетеннями Ауербаха, Мейснера, слизовим та підслизовим сплетеннями.[62]
Черепні нерви (лат. nervi craniales), яких з класичної точки зору налічують 12 пар[52], хоча наявна і 13 пара – термінальний нерв[63], є частиною ПНС, яка відходить від головного мозку. Черепні нерви мають більшу спеціалізацію у порівнянні зі спинномозковими нервами (бувають чутливими, руховими та змішаними). Бувають справжніми (0, III-XII пара) та несправжніми (I-II пара). Останні називаються так тому, що є радше продовженням головного мозку, ніж нервами в класичному розумінні цього слова (при цьому I пара черепних нервів розвивається з однойменної плакоди, а II – є істинним продовженням головного мозку, тому деякі вчені відносять II пару до ЦНС).[52]
Черепні нерви також пов'язані з нервовими вузлами. 0[63], V[64], VII[65], VIII[66], IX[67], X[68] пари черепних нервів мають чутливі вузли, а III[69], VII[70], IX[71] та X[72] пари черепних нервів також пов'язані з парасимпатичними нервовими вузлами.
-
 Будова спинномозкового нерва
Будова спинномозкового нерва -
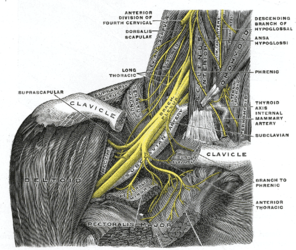 Плечове сплетення
Плечове сплетення -
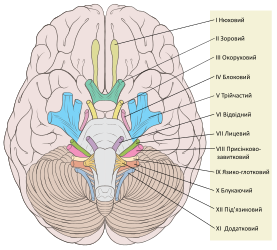 Черепні нерви
Черепні нерви -
 Підслизове (Мейснера) сплетення
Підслизове (Мейснера) сплетення




Синапси та нейромедіатори[ред. | ред. код]

.jpg)
Організація та класифікація[ред. | ред. код]
Місце передачі інформації від нейрона до нейрона називається синапсом[73]. Основна кількість синапсів в нервовій системі тварин – це хімічні синапси[74]. Вони складаються з пре- та постсинаптичної мембран та синаптичної щілини[75].
- Пресинаптична мембрана у більшості випадків є закінченням терміналі аксона[75][76], хоча у випадку дендро-дендричних синапсів представлена поверхнею дендрита[77]. Вона містить велику кількість везикул, у яких містяться молекули нейротрансмітера[75]. У ній також виділяють синаптичну активну зону, де відбувається виділення нейротрансмітера[73].
- Постсинаптична мембрана у випадку міжнейронального синапса може бути представлена дендритом, тілом нейрона, або асконом[75][76]. У випадку нервово-м'язового синапса замість «постсинаптичної мембрани» вживається термін «кінцева пластинка», яка є специфічною ділянкою м'язового волокна[76][78].
- Між цими мембранами розміщена синаптична щілина, у яку виділяється нейромедіатор. Час, якй витрачається на виділення нейромедіатора в синаптичну щілину, його з'єднання з рецептором, викликаня постсинаптичного потенціалу та потенціалу дії, називається синаптичною затримкою.[79].
В хімічному синапсі нервовий імпульс прямує тільки в одному напрямку: від пресинаптичної мембрани до постсинаптичної[73][80].
Хімічні синапси бувають нервово-м'язовими та міжнейрональними[81]. Останні діляться на аксо-дендритні, аксо-соматичні, аксо-аксональні та дендрито-дендричні. Хімічні синапси також діляться в залежності від нейромедіатора, який використовується для передачі збудження.[75]
Нейромедіатори, або нейротрансмітери – це речовини, які забезпечують передачу сигналу від одного нейрона до іншого в хімічному синапсі. Вони бувають різної біохімічної структури (аміни, амінокислоти, нейропептиди).[75][82] За способом дії вони бувають іонотропними, тобто такими, які безпосередньо впливають на проникність іонних каналів та метаботропними – такими, які впливають на іонні канали через посередництво вторинних месенджерів. Перші викликають швидку синаптичну відповідь, а другі – повільну. Іонотропні нейромедіатори взаємодіють з іонотропними рецепторами, які є частинами іонних каналів, а метаботропні нейромедіатори взаємодіють з однойменними рецепторами. Медіатори можуть викликати збудження або гальмування постсинаптичної мембрани. Дію метаботропних нейромедіаторів також називають нейромодуляторною дією, оскільки вони змінюють процеси протікання синаптичної передачі (сприяють або запобігають виникненню потенціалу дії).[83][78]
Окрім хімічних сипапсів існують електричні. В електричному синапсі між двома нейронами існує спеціальний білковий «коридор», через який іони (а отже і струм) та молекули без жодних перешкод прямують в обидві сторони.[74] В деяких тварин вивчені змішані синапси, в яких наявна як пряма електрична, так і хімічна передача імпульса. Відмінною від синаптичної передачі (пряма взаємодія нейронів) є ефаптична (непряма взаємодія нейронів), яка реалізується завдяки близькому розташуванню нервових волокон та дії локальних струмів).
Синаптична передача в хімічному синапсі[ред. | ред. код]

Нейромедіатор в синаптичну щілину виділяється квантами. Один квант – це та кількість молекул нейромедіатора, яка міститься в одній везикулі. Окремі кванти виділяються з пресинаптичної мембрани навіть за відсутності потенціалу дії. Кожен з них провокує виникнення мініатюрного постсинаптичного потенціалу (мініатюрного збуджуючого постсинаптичного потенціалу або мініатюрного інгібуючого потенціалу). Потенціал дії збільшує виділення квантів і разом вони здатні викликати постсинаптичний потенціал.[84][85]
Потенціал дії по нервовому волокну прямує до пресинаптичної мембрани. Тут під його впливом активуються кальцієві потенціалзалежні канали і велика кількість Ca2+ потрапляє в цитоплазму пресинаптичної мембрани. На це реагують чутливі до кальцію білки, які запускають механізм підтягування пухирців до плазматичної мембрани, злиття з нею, і таким чином вміст везикул потрапляє в синаптичну щілину (цей процес називається екзоцитозом).[77][84][86] Нейромедіатор потрапляє в синаптичну щілину, взаємодіє з рецепторомами та викликає постсинаптичні потенціали[79]. До постсинаптичних потенціалів у міжнейрональному синапсі належать збуджувальний постсинаптичний потенціал (ЗПСП) та гальмівний постсинаптичний потенціал (ГПСП), а у нервово-м'язовому синапсі – потенціал кінцевої пластинки (ПКП). Постсинаптичні потенціали не підкоряються закону «все або нічого»: їх величина залежить від сили подразника і вони здатні до сумації. Також при їхньому поширенні вони затихають зі збільшенням відстані і тривають вони довше, ніж потенціал дії.[74][76][87]
Загалом процес синаптичної передачі у цих двох синапсах протікає однаково, проте є дві відмінності, які сильно на нього впливають: нейромедіатор (у нервово-м'язовому синапсі нейромедіатором виступає ацетилхолін, а у міжнейрональному синапсі передача забезпечується великим різноманіттям збуджуючих та гальмівних нейромедіаторів) та амплітуда постсинаптичного потенціалу (ПКП має значно більшу амплітуду, ніж ЗПСП)[81].
У м'язово-нервовому синапсі нервова терміналь має до тисячі активних зон. Потенціал дії спричиняє виділення нейромедіатора в приблизно третині цих зон. На постсинаптичній мембрані міститься велика кількість рецепторів. В сумі ці два фактори спричиняють виникнення ПКП. Амплітуда у ПКП досить велика (прибизно 50 мВ[81]) і достатня для виникнення потенціалу дії у м'язовому волокні, що проявляється скороченням м'яза. Оскільки ПКП в нормі спричиняє потенціал дії, нервово-м'язовий синапс характеризується високою надійністю.[76]
Дещо інакша ситуація наявна в міжнейрональних синапсах. Зазвичай терміналь в такому синапсі має від одної до чотирьох активних зон. Потенціал дії, на відміну від нервово-м'язового синапсу, лише збільшує ймовірність виділення нейромедіатора: на кожен потенціал дії може виділитися одна або дві везикули, а може не виділитися жодної. Окрім того на постсинаптичній мембрані міститься в сто разів менше іонних каналів, ніж на кінцевій пластинці. Тому ЗПСП у міжнейрональному синапсі має маленьку амплітуду (1 мВ і менше[81][88]) і недосягає порогового рівня, після якого виникає потеніал дії.[76] Окрім того, у випадку дії гальмівного медіатора виникає ГПСП і заряд ще більше віддаляється від порогового рівня. Проте, кожен нейрон формує величезну кількість синапсів з іншими нейронами; постсинаптичні потенціали всіх цих нейронів здатні до сумації. Сумація буває часовою, коли один синапс спричиняє декілька постсинаптичних потенціалів підряд, та просторовою, коли сумуються постсинаптичні потенціали багатьох синапсів. Сумуються як ЗПСП, так і ГПСП. Якщо в наслідок сумації досягнуто порогового рівня – виникає потенціал дії в постсинаптичному нейроні.[88][89]
Після дії медіатор або захоплюється назад пресинаптчиною мембраною, або захоплюється астроглією, або дифундує, або деградує під дією фермента[83].
Закони проведення імпульсу в синапсах[ред. | ред. код]
Для хімічних синапсів характерно[80]:
- Однобічність проведення – імпульс передається з пресинаптичної мембрани на постсинаптичну; однак існують двобічні синапси (наприклад, їх дуже багато у кнідарій та реброплавів), по яких імпульси можуть прямувати в обидві сторони;
- Синаптична затримка, яка пов'язана з виділенням нейромедіатора, необхідністю в сумації постсинаптичних потенціалів;
- Вже описана вище сумація
- Трансформація ритму – частота потенціалів дії, які прямують по нервовому волокну до пресинаптичної мембрани, необов'язково викликає таку ж частоту на постсинаптичній мембрані
- Післядія збудження – подовження збудження в ЦНС, пов'язане з відносно довгою тривалістю постсинаптичних потенціалів
- Стомлюваність, яка пов'язана зі зменшенням кількості нейротрансмітера в пресинаптичній мембрані.
З синапсами пов'язані два фундаментальних явища в нервовій системі – збудження та гальмування. І перший, і другий процеси відносяться до активних процесів. Гальмування може бути пресинаптичним (як у випадку аксо-аксонального синапсу, коли аксон гальмівного нейрона закінчується на терміналях аксона іншого нейрона) та постсинаптичним (коли відбується гіперполяризація постсинаптчиної мембрани).[87][90]
Синапси відіграють більшу роль, ніж просто місце передачі імпульсу від нейрона до нейрона. Вони мають великий вплив у процесах пам'яті та навчання завдяки явищу, яке називається синаптична пластичність яке по суті є адаптацією синапса до його використання та навантаження на нього. Синаптична пластичність буває коротко- та довготривалою. Перша пов'язана зі пресинаптичною мембраною, а саме зі збільшенням або зменшенням виділення нейромедіатора, а друга – як з пре-, так і з постсинаптичною мембранами. Це в свою чергу впливає на виникнення потенціалу дії протягом деякого часу (полегшує або ускладнює його виникнення) в постсинаптичному нейроні, що в свою чергу є фізіологічним підгрунтям пам'яті та навчання.[74][91][92]
Неврологія та нейрохірургія[ред. | ред. код]

Наука, яка вивчає порушення у функціонуванні нервової системи (хвороби нервової системи), способи розпізнавання цих порушень та їхнє лікування, називається неврологією. Неврологія є складовою нейронауки. Лікар, який практикується в неврології, відповідно називається неврологом.
Виділяють п'ять груп хвороб нервової системи; уражатися може як ЦНС, так і ПНС, так і обидві частини нервової системи:
- травматичні хвороби – наприклад, забій головного мозку, струс спинного мозку
- пухлинні – наприклад, астроцитома, нейробластома
- судинні – наприклад, геморагічний інсульт, судинна мальформація
- запальні – сюди можна віднести інфекційні хвороби нервової системи (менігококовий менінгіт, наприклад), автоімунні (синдром Гієнна – Барре)
- дегенеративно-дистрофічні, наприклад, бічний аміотрофічний склероз
Для встановлення правильного діагнозу існує неврологічний огляд. Він базується на оцінці різних систем нервової системи. Кожен симптом та їх сукупність (синдром) вказує на стан нервової системи в певному місці, саме тому діагноз в неврології в основному базується на топографії нервової системи (нервових шляхах, локалізації нервових центрів). Однак, топічний діагноз вказує на місце ураження і не зважди вказує на причину та патогенез цього ураження. Тому, окрім опитування та огляду невролог використовує додаткові дослідження – лабораторні та інструментальні. Серед основних інструментальних методів можна виділити ЕЕГ, ЕМГ, КТ, МРТ.[93]
Нейрохірургія теж вивчає захворювання нервової системи, однак, на відміну від консервативного лікування у неврології, застосовує хірургічне лікування – пряме втручання в пошкодженні тканини з метою вилікування. Методи діагностики в нейрохірургії в основному ті ж самі, що у неврології.
Мієлін[ред. | ред. код]

Мієлін є речовиною, багатою на ліпіди. Він покриває аксони нейронів у ЦНС і через свій білий колір утворює білу речовину. Мієлін також покриває аксони, які входять до складу периферичних нервів. При цьому варто зазначити, що є і немієлінізовані аксони. Основна функція мієліну – це прискорення проведення імпульсу по аксону; мієлінізовані волокна передають імпульс в рази швидше у порівнянні з немієлінізованими аксонами. Це пояснюється тим, що мієлін є дуже доброю ізолюючою речовиною. У ЦНС мієлін продукується олігодендроцитами, а у ПНС – шванівськими клітинами.
Потреба в мієліні для ЦНС та ПНС різниться; різниться і біохімічна будова периферійного та центрального мієліну.
Так, для того аби збільшити швидкість передачі імпульсу, необхідно збільшити розмір волокна, що в умовах закритих просторів (черепа та хребта) неможливо. Саме тому, аби не збільшувати об'єм ЦНС, синтезується мієлін (окрім того один олігодендроцит може віддавати свої відростки до великої кількості аксонів). Серед основних білків, притаманних центральному мієліну, головними є основний білок мієліну та ліпофілін та його видозмінена форма DM-20. Відсутність ліпофіліну в ЦНС хребетних спричиняє дегенерацію аксонів.
До складу периферичного мієліну входять в основному периферичний мієліновий білок 22 та мієліновий білок 0.
При різних захворюваннях нервової системи уражається різний мієлін: при розсіяному склерозі страждає центральний мієлін, в той час як периферичний – при хворобі Шарко-Марі-Тута, наприклад.[13][94]
Примітки[ред. | ред. код]
- ↑ Ashwell, 2014, с. 51.
- ↑ Brodal, 2010, с. 5.
- ↑ Головацький, 2 том, 2007, с. 208.
- ↑ Головацький, 3 том, с. 11.
- ↑ Verkhratsky, 2013, с. 76.
- ↑ Verkhratsky, 2013, с. 175.
- ↑ Brodal, 2010, с. 20-22.
- ↑ Squire, 2008, с. 51-52.
- ↑ Verkhratsky, 2013, с. 108.
- ↑ Verkhratsky, 2013, с. 249-250.
- ↑ Verkhratsky, 2013, с. 252.
- ↑ Squire, 2008, с. 48.
- ↑ а б Brodal, 2010, с. 23.
- ↑ Verkhratsky, 2013, с. 322.
- ↑ Verkhratsky, 2013, с. 344-345.
- ↑ Brodal, 2010, с. 26-27.
- ↑ Squire, 2008, с. 53-54.
- ↑ Verkhratsky, 2013, с. 391-392.
- ↑ Verkhratsky, 2013, с. 393.
- ↑ Verkhratsky, 2013, с. 407-408.
- ↑ Verkhratsky, 2013, с. 416-418.
- ↑ Verkhratsky, 2013, с. 420-422.
- ↑ а б в г д е Galizia, 2013, с. 103.
- ↑ а б Squire, 2008, с. 83.
- ↑ Galizia, 2013, с. 104.
- ↑ а б в Squire, 2008, с. 82.
- ↑ а б Galizia, 2013, с. 105.
- ↑ Squire, 2008, с. 80.
- ↑ а б Squire, 2008, с. 32-33.
- ↑ Головацький, 2 том, 2007, с. 203.
- ↑ Головацький, 2 том, 2007, с. 222.
- ↑ а б в Головацький, 2 том, 2007, с. 223.
- ↑ Головацький, 2 том, 2007, с. 224-225.
- ↑ Головацький, 2 том, 2007, с. 226.
- ↑ Головацький, 2 том, 2007, с. 227.
- ↑ Головацький, 2 том, 2007, с. 229.
- ↑ Головацький, 2 том, 2007, с. 224.
- ↑ Головацький, 2 том, 2007, с. 230.
- ↑ Головацький, 2 том, 2007, с. 232.
- ↑ Головацький, 2 том, 2007, с. 233-234.
- ↑ Головацький, 2 том, 2007, с. 237.
- ↑ Головацький, 2 том, 2007, с. 238.
- ↑ Головацький, 2 том, 2007, с. 246.
- ↑ Головацький, 2 том, 2007, с. 248.
- ↑ Головацький, 2 том, 2007, с. 250.
- ↑ Головацький, 2 том, 2007, с. 255.
- ↑ Головацький, 2 том, 2007, с. 252.
- ↑ Головацький, 2 том, 2007, с. 263-266.
- ↑ Головацький, 2 том, 2007, с. 268.
- ↑ Головацький, 2 том, 2007, с. 272.
- ↑ Головацький, 2 том, 2007, с. 274.
- ↑ а б в Головацький, 3 том, 2009, с. 9.
- ↑ Головацький, 3 том, 2009, с. 7.
- ↑ Головацький, 3 том, 2009, с. 41.
- ↑ Головацький, 3 том, 2009, с. 43.
- ↑ Головацький, 3 том, 2009, с. 55.
- ↑ Головацький, 3 том, 2009, с. 57.
- ↑ Головацький, 3 том, 2009, с. 63.
- ↑ Головацький, 3 том, 2009, с. 76.
- ↑ Головацький, 3 том, 2009, с. 82.
- ↑ Головацький, 3 том, 2009, с. 83-86.
- ↑ Головацький, 3 том, 2009, с. 81.
- ↑ а б Головацький, 3 том, 2009, с. 13.
- ↑ Головацький, 3 том, 2009, с. 16.
- ↑ Головацький, 3 том, 2009, с. 25.
- ↑ Головацький, 3 том, 2009, с. 27.
- ↑ Головацький, 3 том, 2009, с. 28.
- ↑ Головацький, 3 том, 2009, с. 30.
- ↑ Головацький, 3 том, 2009, с. 15.
- ↑ Головацький, 3 том, 2009, с. 21.
- ↑ Головацький, 3 том, 2009, с. 22.
- ↑ Головацький, 3 том, 2009, с. 32.
- ↑ а б в Squire, 2008, с. 157.
- ↑ а б в г Brodal, 2010, с. 40.
- ↑ а б в г д е Шевчук, 2012, с. 56.
- ↑ а б в г д е Squire, 2008, с. 160.
- ↑ а б Brodal, 2010, с. 41.
- ↑ а б Squire, 2008, с. 227.
- ↑ а б Шевчук, 2012, с. 57.
- ↑ а б Шевчук, 2012, с. 62-63.
- ↑ а б в г Squire, 2008, с. 228.
- ↑ Squire, 2008, с. 136.
- ↑ а б Brodal, 2010, с. 42-43.
- ↑ а б Brodal, 2010, с. 42-23.
- ↑ Squire, 2008, с. 159.
- ↑ Squire, 2008, с. 161.
- ↑ а б Brodal, 2010, с. 44.
- ↑ а б Brodal, 2010, с. 45.
- ↑ Squire, 2008, с. 242-443.
- ↑ Шевчук, 2012, с. 58-59.
- ↑ Brodal, 2010, с. 51.
- ↑ Squire, 2008, с. 176-179.
- ↑ Гусев, 2009, с. 21.
- ↑ Squire, 2008, с. 49-50.
Для нейрона
Класифікація[ред. | ред. код]

Існує багато класифікацій нейронів, кожна з яких базується на їх морфології, конфігурації відростків, функції, яку останні виконують. Так, відповідно до конфігурації відростків, нейрони бувають[1][2]:
- Уніполярні – нейрони, які широко поширені серед безхребетних тварин, а у хребетних (в тому числі і людини) зустрічаються в основному в період ембріогенезу, хоча, наприклад у людини, їх можна зустріти в корі мозочка (уніполярні щіткові клітини)[3]. Характеризуються тим, що від тіла відходить лише один відросток – аксон. По ходу аксона від нього відходять гілочки, які виконують функції дендритів. В безхребетних вони відходять від аксона в центрі нервового вузла, де утворюють велику кількість синапсів[4].
- Псевдоуніполярні – це ще один специфічний тип нейронів, поширений серед усіх хребетних. Такі нейрони зустрічаються тільки в периферичні нервовій системі, в чутливих вузлах спинномозкових та черепних нервів, які мають чутливі вузли (виключення VIII пара черепних нервів). Їхня особливість полягає в тому, що від тіла відходить лише один відросток – аксон – який ділиться на периферичну гілку, яка виконує функції дендрита (при цьому морфологічно залишається аксоном) і несе сигнал від периферії, та центральну, яка несе сигнал до ЦНС[5].
- Біполярні – нейрони, які мають два відростки: один аксон та один дендрит. Такі клітини можна зустріти в сітківці (біполярні клітини), у вузлах VIII пари черепних нервів[6].
- Мультиполярні – розташовані у ЦНС (пірамідні нейрони, клітини Пуркіньє мозочка) та у вегетативних вузлах ПНС та є переважаючим типом нейронів у хребетних. Мають один аксон та багато нейронів. В залежності від довжини, мультиполярні нейрони ділять на клітини Гольджі I типу, які мають довгі аксони (можуть сягати до 1 м) та клітини Гольджі II типу, які мають дуже корокі, довжиною в декілька мікрометрів, аксони.
За функціональною класифікацією нейрони можуть бути:
- аферентним нейроном, або протонейроном, або чутливими нейроном — це той нейрон, який першим сприймає подразнення з периферії і він несе його в напрямку ЦНС; такі нейрони розташовані за межами ЦНС (виняток — нейрони середньомозкового ядра трійчастого нерва). Загалом такими нейронами є псевдоуніполярні нейрони: їх периферична гілка аксона закінчується рецептором, а центральна — синапсом.[7]
- інтернейроном, або дейтеронейроном – це той нейрон, який передає сигнал від одного нейрона до іншого. І дендрит, і аксон такого нейрона мають синапс. Такі нейрони складають абсолютну більшість нейронів нервової системи.[8]
- моторним нейроном, або мотонейроном — це той нейрон, який безпосередньо іннервує орган чи м'яз (відповідно є соматомоторні та вісцеромоторні нейрони). Розташовані такі нейрони як в ЦНС, так і в ПНС.[9]
Примітки2[ред. | ред. код]
- ↑ Головацький, 2 том, 2007, с. 204.
- ↑ Ganong, 2012, с. 85.
- ↑ Enrico Mugnaini, Gabriella Sekerková, Marco Martina (2011). The unipolar brush cell: A remarkable neuron finally receiving deserved attention. Brain Research Reviews. 66 (1-2): 220—245. doi:10.1016/j.brainresrev.2010.10.001. PMID 20937306.(англ.)
- ↑ Squire, 2008, с. 21.
- ↑ Brodal, 2010, с. 5.
- ↑ Brodal, 2010, с. 6.
- ↑ Squire, 2008, с. 17.
- ↑ Squire, 2008, с. 19-20.
- ↑ Squire, 2008, с. 18.