Поле Бродмана 45
| Поле Бродмана 45 | |
|---|---|
 Бічна поверхня лівої півкулі головного мозку (трикутна ділянка показана жовтим) | |
 | |
 | |
| Деталі | |
| Ідентифікатори | |
| Латина | Area triangularis |
| NeuroLex ID | birnlex_1777 |
| FMA | 68642 |
| Анатомічна термінологія | |
Поле Бродмана 45 (BA45), є частиною лобової кори в мозку людини. Розташоване на бічній поверхні, нижче поля Бродмана 9 (BA9) і прилягає впритул до поля Бродмана 46 (BA46).
Це поле також відоме як трикутна ділянка (нижньої лобової звивини), й знаходиться навколо передньої горизонтальної гілки латеральної борозни, орбітальної частини нижньої лобової звивини. Каудально поле оточене передньою висхідною гілкою латеральної борозни , межує з острівцем в глибині бічної борозни.
З точки зору цитоархитектоніки, поле обмежене каудально полем Бродмана 44 (BA44), ростродорзально межує з полем Бродмана 46 (BA46), і вентрально з орбітальним полем Бродмана 47 (BA47) (Корбініан Бродман, 1909).
Функції[ред. | ред. код]
Разом з BA 44, поле містить Центр Брока, ділянку, яка активно бере участь у виконанні семантичних завдань, таких як смислове рішення задачі (визначення, чи є слово абстрактним чи конкретним поняттям) чи завдань на ув'язування дієслів з іменниками).
Точна роль BA45 у виконанні семантичних завдань залишається предметом дискусій. Для деяких дослідників, його роль полягає в сприянні семантичному пошуку або семантичній робочій пам'яті. Згідно з цією думкою, BA44 і BA45 разом сприяють відновленню семантичної інформації та оцінці відновленої інформації з урахуванням критеріїв, відповідних для даного контексту.[1][2] дещо змінений варіант цієї думки полягає в тому, що активація BA45 потребується лише при контрольованому семантичному пошуку, коли сильні стимул-стимул асоціації відсутні.[3] Для інших дослідників роль BA45 не обмежується семантикою як такою, але включає й роботу в усіх видах діяльності, що вимагають рішень, відповідних до завдань з числа "конкуруючих" рішень.[4] Ураження BA45 приводять до характерних симптомів експресивної афазії у людей, позбавлених півкульної домінанти.
Результати досліджень[ред. | ред. код]
Асиметрія і мовна домінанта[ред. | ред. код]

Потужна кореляція була виявлена між мовою-мовленням та анатомічно асиметричною трикутною ділянкою. Фундес (Foundas), і співавт. показали, що мовні функції можуть бути локалізовані в одній області мозку, (як і показав попередньо, Поль Брока), але ними була висловлена ідея про те, що одна сторона мозку більш тісно пов'язана з мовою, ніж інша. Людський мозок має дві півкулі, і кожна з них схожа на іншу; тобто, ніби то, одна півкуля повинна являти собою дзеркальне зображення іншої. Насправді це не так. Foundas, і співавт. виявили, що ліва частина зони Брока а саме, трикутна ділянка (лат. pars triangularis), насправді більше, ніж така сама ділянка, в правій півкулі мозку. Цікаво, що ця "лівобічна асиметрія" - як анатомічна, так і функціональна. Це означає, що частина мозку, яка активна під час обробки мовної інформації - за розміром більша. Майже у всіх обстежених це була ліва сторона. По суті, єдиний з обстежених, у якого було виявлене правопівкульне домінування мови, мав правобічну асиметрію цієї трикутної ділянки.[5]
Деякі інші дослідники, проте, не знайшли об'ємної асиметрії трикутної ділянки. Вони оскаржували попередні повідомлення й припустили, що суперечності в попередніх висновках можуть бути пов'язані з великою мінливістю морфології трикутної ділянки. Тобто, у різних людей трикутні ділянки, як правило, розрізняються за розміром і формою набагато більше, ніж інші ділянки мозку, такі як, наприклад, глибокі кіркові ядра. Крім того, хоча ці дослідника знайшли статистично значиму асиметрю в pars opercularis та planum temporale[en], вони не знайшли жодної кореляції між асиметрією цих ділянках мозку з розмірами й асиметрією трикутної ділянки.[6]
Зв'язки в нижній лобовій звивині[ред. | ред. код]
Принаймні, одне дослідження виявило високий ступінь зв'язку між трьома підрозділами нижньої лобової звивини (gyrus frontalis inferior, inferior frontal gyrus (IFG)). З допомогою стимуляції однієї області IFG і вимірювання відповіді в різних ділянках дослідники змогли показати існування численних провідних шляхів між трикутною ділянкою й ділянкою кришечки. [7]
Крім того, трикутна ділянка залучена до семантичної обробки мови. Шляхом дослідження за допомогою електроенцефалографії, відповіді мозку на різні типи речень (зі смисловими помилками й без), Maess і співавтори демонстрували часове відставання при осмисленні помилкових речень. Щоб зрозуміти це відставання, варто лише уявити собі людину, котрій сказали щось незрозуміле. Така людина зупиняється й прагне скористатися моментом, щоби осмислити інформацію. Крім того, дослідники продемонстрували характерний енцефалографічний патерн, який називається “N400” й відповідає негативній реакції трикутної ділянки через приблизно 400 мс після синтаксично невідповідного подразника.[8] Однак, трикутна ділянка, швидше за все, є лише частиною мережі, яка генерує патерн “N400” на ЕЕГ, бо магнітний аналог N400m, досліджений за допомогою МЕГ (магнітоенцефалографії) незмінно локалізується у верхній скроневій корі.[9]
Префронтальна кора і когнітивний контроль пам'яті[ред. | ред. код]
Трикутна ділянка, як виявилося, грає певну роль в когнітивному контролі пам'яті.[10]
При читанні вголос, людина повинна розшифрувати написане розшифрувати його вимову. Ця обробка відбувається в центрі Брока. Читач може використати попередні знання слова для того, щоб правильно вимовити його, або читач може використовувати знання з систематичного поєднання букв, які представляють відповідні фонеми. Вчені можуть дізнатися про те, що робить мозок, слідкуючи за помилками в мові. Як зазначено вище, вчені можуть досліджувати додаткову роботу мозку, коли люди стикаються з проблемою. У цьому випадку, вчені скористалися способом псевдо-слів і слів-виключень та дослідженням пацієнтом інтерпретації цих проблемних слів. Протягом обробки мовної інформації в мозку людини використовуються різні частини центру Брока для різних речей. Трикутна ділянка' бере участь у певному типі обробки мовної інформації. Зокрема, трикутна ділянка активізується, коли люди читають слова-виключення.
Кожна людина у світі вивчила принаймні одну мову. Також, майже всі, що вивчила мову, зробили це в молодому віці. Деякі люди багатомовні. Деякі з цих багатомовних навчилися другої або третьої мови з позицій своєї основної (першої мови) у молодому віці, а деякі і зовсім вивчили інші мови в зрілому віці. Дослідження різних піднаборів одномовних і багатомовних осіб показали деякі цікаві результати.
Дивлячись на схожість між першою і другою мовою і процесами, що виникають при цьому в мозку, ці дослідники виявили, що активація мозку виглядає зовсім по-іншому в залежності від того, якою мовою оперували піддослідні особи. [11]
Це дослідження показує, що, врешті, білінгви використовують трикутну ділянку набагато більше, ніж їхні колеги. Відповідно до цієї точки зору, носії мови можуть говорити досить швидко, в той час як їхні пізньо-двомовні колеги (ті, хто вивчили мову в зрілому віці) змушені затинатися в процесі обробки граматичних правил. [12]
Динаміка кіркового розпізнавання слів[ред. | ред. код]
Існує теорія, згідно з якою, зокрема, трикутна ділянка особливо залучена до семантичної обробки мови, на відміну від фонологічної обробки. [13]
Інше дослідження називалося дослівно: "Семантичне кодування і пошук у лівій нижній префронтальній корі головного мозку: функціональнамагнітно-резонансна томографія дослідження складності завдань та специфіка процесу." Ці дослідники виявили, що трикутна ділянка (а також деякі сусідні) збільшує свою активність протягом семантичного кодування, при чому, незалежно від складності оброблюваного слова. Це узгоджується з теорією, яка говорить, що трикутна ділянка бере участь у семантичної обробки більш ніж у фонологічній обробці. Крім того, вони виявили, що це семантичне кодування призводять до зменшення участі трикутної ділянки в повторенні вживаних слів. [14]
Центр Брока, головний мозок: нові рамки[ред. | ред. код]
У людини регіони мозку тісно взаємопов'язані між собою, собливо в лівій лобової мовній мережі. Мова може бути інтегрованою в багато, здавалося б, не пов'язаних між собою розумових процесів. Насправді, це було б малоймовірно для трикутної ділянки не мати кілька ролей у головному мозку, особливо враховуючи його високий ступінь зв'язності з іншими ділянками. [15]
Шизофренія та центр Брока[ред. | ред. код]
Шизофренія - це малозрозуміле, й донині, захворювання зі складними симптомами. У спробі знайти його причину, дослідники неодноразово досліджували мозок хворих. Раніше було показано, що у пацієнтів із шизофренією виникають аномальні малюнки борозен і звивин у мозку, його асиметрія, складність і мінливість. Ці дослідники представили дані, що показують, що трикутна ділянка у хворих шизофренією була сильно спотворена порівняно зі середньостатистично нормальними особами. Дослідники стверджували, що центр Брока - особливо пластична ділянка й його морфологія може різко змінюватися від дитинства до дорослого віку. Це пояснює здібності дітей до легкого вивчення мов, але це також означає, що залучення центру Брока обмежене по відношенню до пам'яті і спогадів; діти, схоже, не можуть усвідомлено шукати свої спогади. Крім того, дослідники взяли заміри сірої і білої речовини мозку піддослідних і порівняли ці значення з контрольною групою. Вони виявили, що у хворих шизофренією різко редукується біла речовина.
По мірі розвитку мозку з віком, зв'язки різних регіонів мозку зазнають різких змін. Дослідники виявили, що існує диспропорція розвитку білої й сірої речовини у хворих на шизофренію. У шизофреніків, як правило, помітна відсутність експансії білої речовини.[16]
Евристичий аналіз та аналітичне мислення[ред. | ред. код]
Транскраніальна магнітна стимуляція застосована до лівої BA45 виявила неконгруентну продуктивність розумових процесів.[17]
Зображення[ред. | ред. код]
-
 Вид спереду.
Вид спереду. -
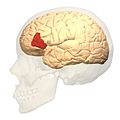 Боковий вигляд.
Боковий вигляд. -
 Бічна поверхня лівої півкулі головного мозку, при погляді збоку. (показано помаранчевим кольором).
Бічна поверхня лівої півкулі головного мозку, при погляді збоку. (показано помаранчевим кольором).


Посилання[ред. | ред. код]
- ↑ Gabrieli та ін. (1998). The role of left prefrontal cortex in language and memory. PNAS. 95 (3): 906—913. doi:10.1073/pnas.95.3.906. PMC 33815. PMID 9448258.
- ↑ Buckner, R. (1996). Contributions of specific prefrontal brain areas to long-term memory retrieval. Psychonomic Bulletin and Review. 3 (2): 149—158. doi:10.3758/BF03212413.
- ↑ Wagner, A. D. (2002).
- ↑ Thompson-Schill та ін. (1999). Effects of repetition and competition on activity of left prefrontal cortex during word generation. Neuron (journal). 23 (3): 513—522. doi:10.1016/S0896-6273(00)80804-1. PMID 10433263.
- ↑ Foundas AL, Leonard CM, Gilmore RL, Fennell EB, Heilman KM (January 1996). Pars triangularis asymmetry and language dominance. Proc Natl Acad Sci U S A. 93 (2): 719—722. doi:10.1073/pnas.93.2.719. PMC 40120. PMID 8570622.
- ↑ Keller, SS; Highley, JR; Garcia-Finana, M; Sluming, V; Rezaie, R; Roberts, N (2007). Sulcal variability, stereological measurement and asymmetry of Broca's area on MRI images. J Anat. 211 (4): 534—55. doi:10.1111/j.1469-7580.2007.00793.x. PMC 2375829. PMID 17727624.
- ↑ Jeremy D.W. Greenlee; Hiroyuki Oya; Hiroto Kawasaki; Igor O. Volkov; Meryl A. Severson III; Matthew A. Howard III; John F. Brugge (2007). Functional connections within the human inferior frontal gyrus. The Journal of Comparative Neurology. 503: 550—559. doi:10.1002/cne.21405. PMID 17534935.
- ↑ Maess, Burkhard; Herrmann, Christoph S.; Hahne, Anja; Nakamura, Akinori; Friederici, Angela D. (2006). Localizing the distributed language network responsible for the N400 measured by MEG during auditory sentence processing. Brain Research. 1096 (1): 163—172. doi:10.1016/j.brainres.2006.04.037. PMID 16769041.
- ↑ Vartiainen, J; Parviainen, T; Salmelin, R (2009). Spatiotemporal convergence of semantic processing in reading and speech perception. Journal of Neuroscience. 29: 9271—9280. doi:10.1523/jneurosci.5860-08.2009. PMID 19625517.
- ↑ David Badre & Anthony D. Wagner (2007). Left ventrolateral prefrontal cortex and the cognitive control of memory. Neuropsychologia. 45 (13): 2883—2901. doi:10.1016/j.neuropsychologia.2007.06.015. PMID 17675110.
- ↑ H. Jeong, M. Sugiura, Y. Sassa, T. Haji, N. Usui, M. Taira, K. Horie, S. Sato, R. Kawashima.
- ↑ S. okoyama, H. Okamoto, T. Miyamoto, K. Yoshimoto, J. Kim, K. Iwata, H. Jeong, S. Uchida, N. Ikuta, Y. Sassa, W. Nakamura, K. Horie, S. Sato, and R. Kawashima.
- ↑ N. Mainy, J. Jung, M. Baciu, P. Kahane, B. Schoendorff, L. Minotti, D. Hoffmann, O. Bertrand, J. Lachaux.
- ↑ Demb, J.; Desmond, J.; Wagner, A.; Vaidya, C.; Glover, G.; Gabrieli, J. (1995). Cortex: A Functional magnetic resonance imaging Study of Task Difficulty and Process Specificity. The Journal of Neuroscience. 15: 5870—5878.
- ↑ P. Hagoort.
- ↑ J. Wisco, G. Kuperberg, D. Manoach, B. Quinn, E. Busa, B. Fishl, S. Heckers, and A. Sorensen.
- ↑ Tsujii, Takeo; Masuda, Sayako; Akiyama, Takekazu; Watanabe, Shigeru (2010). The role of inferior frontal cortex in belief-bias reasoning: an rTMS study. Neuropsychologia. 48 (7): 2005—2008. doi:10.1016/j.neuropsychologia.2010.03.021.
{{cite journal}}: Cite має пустий невідомий параметр:|1=(довідка)
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||