Поле Бродмана 10
| Поле Бродмана 10 | |
|---|---|
 | |
 | |
| Деталі | |
| Ідентифікатори | |
| Латина | Area frontopolaris |
| NeuroNames | 76 |
| NeuroLex ID | birnlex_1741 |
| FMA | 68607 |
| Анатомічна термінологія | |
Поле Бродмана 10 (BA10) — одне з визначених Корбініаном Бродманом цитологічно визначених полів кори головного мозку. Це передня частина префронтальної кори в мозку людини.[1] BA10 було спочатку визначене Корбініаном Бродманом лише приблизно, виходячи з відмінностей цитоархітектоніки, які були помічені в трупних препаратах. Але з огляду на те, що навіть сучасні методики функціональної візуалізації також не можуть точно визначити межі поля, питання про його кордони залишається відкритим.
BA10 — найбільше з існуючих полів кори в людському мозку. Воно було описане як «одне з найменш зрозумілих регіонів людського мозку». дослідження припускає, що поле бере участь у стратегічних процесах пам'яті й різних виконавчих функціях. В ході еволюції, функції цієї ділянки в результаті розширилися по відношенню до іншої частини мозку.
Анатомія[ред. | ред. код]
Розмір[ред. | ред. код]
Обсяг BA10-близько 14 см3 і становить приблизно 1,2 % від загального обсягу об'єму мозку. Це вдвічі більше, ніж у людиноподібних з однаковим з людиною обсягом мозку. Для порівняння, обсяг BA10 у карликового шимпанзе становить близько 2,8 см3, і це лише 0,74 % від об'єму його мозку.
В кожній півкулі, площа 10 містить приблизно 250 мільйонів нейронів.
Розташування[ред. | ред. код]
BA10 є частиною визначеної цитоархітектонічно лобової ділянки кори головного мозку. Поле займає найбільш ростральну частину верхньої лобової звивини і середньої лобової звивини. В людському мозку, у медіальній частині півкулі вона обмежена знизу верхньою ростральною борозною. Вона не поширюється до поясної борозни. Цитоархітектонічно межує дорсально із полем Бродмана 9, каудально з полем Бродмана 46, а знизу з орбітальним полем Бродмана 47 і полем Бродмана 12.
Цитоархитектоніка[ред. | ред. код]
У шість шарів кори головного мозку в районі 10 були описані як такі, що виглядають «на диво, гомогенними». Всі вони легко ідентифікуються. По відношенню один до одного, шар І тонкий середній завширшки, займає становить 11 % від товщини поля 10. Другий шар тонкий і містить дрібні зернисті і пірамідальної середньо-темного забарвлення клітини при фарбуванні по Нісслю, яке зафарбовує РНК і ДНК. Найширший шар третій. Його пірамідальних нейронів менше, розташовані вони ближче один до одного, ніж у вищого шару II й нижчого шару IV. Шари II і III складають 43 % товщі кори. Шар IV тонкий і має чіткі межі з сусідніми шарами III і V . Його клітини бліді, середнього зафарбування. Шар V широкий та складається з двох окремих підшарів, Va і Vb. Щільність клітин Va більше, ніж в Vb, підшар Va має й темніше забарвлення. Шари IV і V складають 40 % товщини кори. Шар VI містить темні пірамідні і веретеноподібні нейрони. Він складає 6 % товщини Поля 10.
Поле 10 відрізняється від сусіднього Поля Бродмана 9 тим, що останній має більш виражений шар Vb і більш помітний шар II. Сусіднє Поле 11 Бродмана порівняно з Полем 10 має більш тонкий шар IV з більш помітними шарами Va, Vb і II.
Поле 10 у людини має найнижчу щільність нейронів серед приматів. Також, його нейрони мають особливо великі дендритні розгалуження дуже щільно розташовані дендритні шипики. Вважається, що це уможливлює інтеграцію вхідної інформації з різних ділянок.
Структура поля[ред. | ред. код]
BA 10 розділене на три підполя, 10p, 10m і 10r. Підполе 10р займає фронтальний полюс, в той час як два інших накривають вентромедіальну частину префронтальної кори. Підполе 10м має тонкі шари II і IV і більш помітний шаром V. На противагу цьому, підполе 10р має помітний шар II і більш товстий шар IV. Великі пірамідальні клітини також присутні в підполі 10р. Шар III також великий. Але відзначається, що «відмінності між трьома підполями змазані й важко провести різкі межі між ними».
Зв'язки[ред. | ред. код]
Дослідження на приматах показали, що поле 10 контактує з іншми вищими асоціативнми ділянками кори, отримуючи вхідну й посилаючи вихідну інформацію. Зокрема, йдеться про префронтальну кору, первинну сенсорну й моторну ділянки. Через крайню капсулу поле пов'язане волокнами зі слуховою та мультисенсорною ділянками верхньої скроневої борозни. Волокна також продовжуються у медіальний поздовжній пучок, у білу речовинуі верхньої скроневої звивини (зони TAa, TS2, і TS3) і найближчі мультисенсорні зони на верхньому краї верхньої скроневої борозни (TPO). Ще одна область, пов'язана з цим полем через крайню капсулу - це вентральна ділянка острівцевої кори. Волокна до поясного пучка поєднують Поле 10 із передньою поясною корою, задньою поясною корою, і ретроспленальною корою (Поле Бродмана 26, Поле Бродмана 29 і Поле Бродмана 30). Гачкуватий пучок з'єднує його з мигдалеподібним тілом, темпорополярною корою і передньою частиною верхньої скроневої звивини. Поле не має зв'язків з тім'яною корою, потиличою корою, інтеротемпоральною корою.
В певних дослідженнях відзначено, що Поле 10 може бути анатомічною структурою для формування абстрактної обробки інформації та інтеграції результатів когнітивних операцій.
Еволюція[ред. | ред. код]
Катерина Семендефері (Katerina Semendeferi) зі співавторами вважає, що «в ході еволюції Поле 10 пройшло пару .. змін: одна включає в себе суттєве збільшення загального розміру поля, а інша - певне збільшення зв'язків особливо з іншими вищими асоціативними полями».
Дослідження рельєфу всередині черепа у Homo floresiensis виявило зростання в ділянці лобового полюсу, що може говорити про збільшення поля 10 Бродмана.
Функція[ред. | ред. код]
Хоча ця ділянка в людському мозку й велика, її функція є маловивченою. Koechlin & Hyafil запропонували вважати основною функцією фронтополярної кори когнітивний розподіл.[2] , що дозволяє раніше запущені завдання переводити у стані очікування для подальшої актуалізації й виконання після закінчення поточного завдання.
Багато які з наших складних форм поведінки і психічної діяльності вимагають одночасної участі декількох завдань. Дослідники припускають, що передня префронтальна кора може виконувати предметно-загальні функції плануванні складних паралельно-послідовних операцій. .[3] У 2006 році Мета-аналіз показав, що ростральна префронтальна кора задіяна в процесах робочої пам'яті, епізодичної пам'яті й координації паралельно-послідовного виконання завдань.[4]
Зображення[ред. | ред. код]
-
 Анімація.
Анімація. -
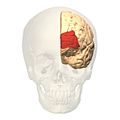 Фронтальний вигляд..
Фронтальний вигляд.. -
 Латеральний вигляд.
Латеральний вигляд. -
 Медіальний вигляд.
Медіальний вигляд.




Див. також[ред. | ред. код]
Примітки[ред. | ред. код]
- ↑ Knowlton, Barbara J.; Morrison, Robert G.; Hummel, John E.; Holyoak, Keith J. (July 2012). A neurocomputational system for relational reasoning. Trends in Cognitive Sciences. 16 (7): 373—381. doi:10.1016/j.tics.2012.06.002. PMID 22717468.
- ↑ Koechlin, E.; Hyafil, A. (2007). Anterior prefrontal function and the limits of human-decision making. Science. 318: 594—598. doi:10.1126/science.1142995.
- ↑ Burgess, P.W.; Dumontheil, I.; Gilbert, S.J. (2007). The gateway hypothesis of rostral prefrontal cortex (area 10) function. Trends in Cognitive Science. 11: 7.
- ↑ Gilbert, Sam J.; Spengler, Stephanie; Simons, Jon S.; Steele, J. Douglas; Lawrie, Stephen M.; Frith, Christopher D.; Burgess, Paul W. (1 червня 2006). Functional specialization within rostral prefrontal cortex (area 10): a meta-analysis. Journal of Cognitive Neuroscience. 18 (6): 932—948. doi:10.1162/jocn.2006.18.6.932. ISSN 0898-929X. PMID 16839301.
Посилання[ред. | ред. код]
- Поле Бродмана 10
- BrainMaps, Поле Бродмана 10 Префронтальної Кори
- Поле Бродмана 10, ключ до еволюції людини?
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||