У цьому термінологічному словнику пояснюються технічні терміни, які зазвичай використовуються при описі скам'янілостей тіл динозаврів. Окрім специфічних термінів, що стосуються динозаврів, він охоплює терміни ширшого вжитку, якщо вони мають центральне значення у вивченні динозаврів або якщо їхнє обговорення в контексті динозаврів є корисним. Глосарій не охоплює іхнологічні чи гістологічні терміни, а також вимірювальні величини.
{{якір|акроміон}}{{term|акроміон}}
{{defn|(лат.acromion) — кістковий гребінь на зовнішньому дистальному кінці лопатки, що забезпечує прикріплення ключиці[1]. У нодозаврид розвивалася яскраво виражена шпора, відома як псевдоакроміон, яка, ймовірно, була місцем прикріплення переднього плечового м'яза (лат.musculus scapulohumeralis anterior), а отже, була аналогом акроміона ссавців[2].}}
аксонія (лат.axony) — розташування домінантного (найбільшого і найбільш виступаючого) пальця на руці або нозі, і найчастіше використовується для опису слідів ніг. Коли центральний палець (як правило, палець III) є домінуючим, як у випадку з лапами більшості двоногих динозаврів, кисть або стопа є мезаксонічною. Ентаксонічна кисть або стопа має медіальний домінантний палець (палець I, або II); цей рідкісний стан можна зустріти у завроподів. І навпаки, ектаксонічна кисть/стопа має домінуючий палець назовні (найчастіше це палець IV). Коли пальців два або чотири й жодна з половин кісті/стопи не є домінантною — такий стан називається параксонічним[3].
арктометатарсус (лат.arctometatarsus, від «стиснутий») — особливість будови стопи, де дві крайні плеснові кістки дуже сильно затискають середню, утворюючи майже єдину структуру. Ця ознака зустрічається у деяких похідних тероподів, включаючи орнітомімових, тиранозаврових, троодонтид, ельмізаврових та авімімових. Існує гіпотеза, що ця адаптація грала роль в амортизації бігу, що було важливим для активних хижаків[4][5]. Спочатку ця особливість використовувалася для визначення клади Arctometatarsalia, яка зараз вважається поліфілетичною[6].
атлас, атлант (лат.atlas) — перший (передній) хребецьхребетного стовпа. Він приймає потиличний відросток основи черепа, утворюючи таким чином з'єднання між черепом і хребтом. Динозаври мали один виросток (лат.condylus), що дозволяв здійснювати обертальний рух між атлантом і черепом. У ссавців, з іншого боку, спостерігаються подвійні виростки, що обмежують рух в одній площині — обертальний рух відбувається переважно між атлантом і другим, осьовим хребцем. Атлант динозаврів був примітивним, та складався з трьох окремих елементів: неврапофіза атланта (англ.atlantal neurapophysis), у верхній частині хребця[7], інтерцентра (лат.intercentrum) спереду та тіла хребця (лат.centrum) ззаду. Додаткова маленька кістка, проатлант, розташована перед невральною дугою (англ.neural arch)[8]. Центральну частину атланта також називають одонтоїдом (лат.odontoid). У динозаврів ці три елементи зазвичай залишаються окремими, але можуть зростатися між собою. У цератопсів атлант, а також осьовий і третій хребець зростаються в єдину кістку[8].
базальні бугри потилиці (лат.basal tubera) — пара горбкоподібних розширень на нижньому боці черепної коробки, які функціонують як місця прикріплення нижніх м'язів шиї. Вони утворені основною потиличною (лат.basioccipitale) та основною клиноподібною (лат.basisphenoid) кістками та сильно виражені у ящеротазових, але лише помірно — у птахотазових[9][10].
базікраній (лат.basicranium) — дно черепної коробки, утворене основною потиличною, основною клиноподібною кістками та парасфеноїдом[11].
бічна клиноподібна кістка (англ.laterosphenoid або pleurosphenoid) — крилоподібна кістка передньої частини бічної стінки черепної коробки. Не видима ззовні, вона розташована спереду від передньовушної й контактує з тім'яною, лобовою та заочною кістками, розташованими зверху. Бічна клиноподібна як окрема кістка є унікальною для архозаврів; в інших рептилій ця частина хряща не закостеніла[12][9][13][14].
бічні потиличні кістки (англ.exoccipital) — парні кістки, що разом з непарними верхньою потиличною та основною потиличною кістками утворює потилицю в задній частині черепа[13][12].
велика гомілкова кістка (англ.tibia) — більша з двох кісток гомілки. Вона розташована медіально від малогомілкової кістки. Великогомілкова кістка має помітний гребінь, що відходить від її передньої поверхні на верхньому кінці — великогомілковий гребінь або кнеміальний гребінь (англ.cnemial crest). Великогомілковий гребінь фіксує сухожилля триголового м'яза гомілки, яке відповідає за розгинання коліна та підсилює ці м'язи[2][15].
верхньощелепова кістка (англ.maxilla) — задня і, як правило, найбільша зубоносна кістка верхньої щелепи. Спереду вона з'єднується з передщелепною і часто формує межі зовнішнього носового отвору. На більшій частині своєї довжини верхня щелепа утворює вентральну межу передочного отвору (англ.antorbital fenestra), а також заглиблення навколо цього отвору — передочну ямку (англ.antorbital fossa), глибоко занурену в верхню щелепу. У похідних тероподів верхня щелепа може мати два менших додаткових отвори — верхньощелепний та передщелепний отвори. На задньому кінці верхня щелепа зчленовується з виличною та слізною кістками[12][15].
верхня потилична кістка (англ.supraoccipital) — непарна кістка потилиці в задній частині черепа. Вона утворює верхній край великого потиличного отвору (лат.foramen magnum) і з'єднується дорсально з тім'яними кістками. На ньому зазвичай можна побачити серединний гребінь, на якому закріплюються м'язи шиї[13][12].
вилична кістка або скулова кістка (англ.jugal) — кістка черепа, яка окреслює нижню межу очної ямки та з'єднується спереду з верхньою щелепою, а ззаду — з квадратно-виличною. Її задній кінець роздвоюється в місці зчленування з квадратно-виличною, що вважається синапоморфією динозаврів[15][12].
відростки хребців (англ.neural spines або spinous processes) — відростки, що підіймаються від верхньої поверхні невральної дуги (англ.neural arch). Утворюють єдиний ряд по середній лінії хребта, причому їхні кінчики зазвичай є вищими точками окремих хребців. Відростки хребців забезпечують поверхні прикріплення для м'язів і зв'язок, що йдуть поверх хребта[16]. У деяких видів вони можуть бути сильно витягнуті, утворюючи «вітрило», або глибоко роздвоєні, так що їхні верхівки утворюють подвійний ряд гребенів[17][18].
вінцевий відросток Вінцевий відросток нижньої щелепи (англ.coronoid process) — кістковий виступ, який тягнеться вгору від верхньої поверхні нижньої щелепи за зубним рядом. Цей відросток служить місцем прикріплення м'язів, які змикають щелепу; вищий вінцевий відросток збільшує плече важеля цих м'язів, а отже, і силу прикусу[19][12].
вторинне піднебіння (англ.secondary palate) — дахоподібна структура, що відокремлює носові дихальні шляхи від рота. Хоани примітивних рептилій відкриваються в ротову порожнину в передній частині рота, що дозволяло блокування дихальних шляхів їжею під час годівлі. Вторинне піднебіння, як це зазвичай буває у ссавців, зміщує хоани назад, дозволяючи харчуватися і дихати одночасно[20]. У деяких кладах, включаючи анкілозаврів[21], спінозаврів[22] і маніраптоподібнихцелурозаврів[23], вторинне піднебіння було кістковим і називалося кістковим вторинним піднебінням (англ.osseous secondary palate). Водночас м'яке вторинне піднебіння було виявлено у деяких інших таксонів, таких як Plateosaurus і Stegosauria, на основі виступів і гребенів піднебінних кісток[24][25]. Кісткове вторинне піднебіння зазвичай утворене полицеподібними розширеннями передщелепних та верхньощелепних кісток, які з'єднуються з лемішем на серединній лінії черепа. Анкілозаври розвинули нову додаткову частину вторинного піднебіння, яка простягалася вперед від крилоподібної та піднебінної кістки в задній частині піднебіння, і має назву каудовентрально піднебінна полиця (англ.caudoventral palatal shelf) (відповідно, передня частина, утворена передщелепною та верхньощелепною кістками, називається ростродорсальною піднебінною полицею (англ.rostrodorsal palatal shelf))[21].
ґленоїд. Суглобова поверхня, утворена коракоїдом і лопаткою, що приймає головку плечової кістки, утворюючи плечовий суглоб. Ґленоїд (лат.glenoid) — увігнута кругла структура, облямована кістковим обідком і розташована на задній поверхні коракоїда та задньодистальній поверхні лопатки[26][15].
дельтопекторальний гребінь. Направлений вперед кістковий виріст на верхній частині плечової кістки. Особливо довгий і помітний дельтопекторальний гребінь (англ.deltopectoral crest) є синапоморфією динозаврів, тобто ознакою, що відрізняє групу від інших груп. У динозаврів гребінь становить 30—40 % довжини плечової кістки. Він забезпечував поверхні для м'язів плеча і грудей (дельтовидних і грудних м'язів відповідно), які, скорочуючись, притягували руку до тулуба[12].
дзьобоподібна кістка, коракоїдна, вороняча кістка або коракоїд (англ.coracoid) — парна кістка грудного пояса, яка кріпиться до нижнього кінця лопатки. Цей плаский, а у динозаврів, як правило, напівкруглий або квадратний за формою елемент, має ендохондральне походження. Його нижній край, разом з лопаткою, утворює частину ґленоїда плечового суглоба або суглобову западину лопатки (англ.glenoid fossa)[12]. На бічній поверхні коракоїда є отвір — коракоїдний отвір — через який проходить надкоракоїдний нерв[26].
додаткова надочноямкова кістка (англ.palpebral, supraorbital або adlacrimal) — невелика кістка, прикріплена до верхнього краю очниці (очної ямки) або утворює її частину. Вона формується у верхній повіці та походить з остеодерми. З обох боків черепа може бути одна або кілька додаткових надочноямкових кісток. Наявність palpebral є синапоморфієюптахотазових, хоча вони також розвинулися в інших кладах рептилій, включаючи крокодилоподібних[27].
ендокран. Зліпки мозкових порожнин, включаючи черепну коробку та невральні канали хребців. Таким чином, ендокрани (англ.endocast можуть фіксувати зовнішні ознаки мозкових структур, які були присутні в цих порожнинах, а насамперед — головного мозку. Ендокрани не є точними копіями, оскільки мозкові порожнини, як правило, містять додаткову тканину, яка може ускладнювати морфологію мозкової структури. Природний ендокран утворюється, коли мозкова порожнина заповнюється осадовими відкладеннями, в той час як штучний ендокран можна виготовити за допомогою ливарного матеріалу[28].
епіосіфікації Додаткові закостеніння (англ.epiossifications), що росли на черепах цератопсів[29]. До них відносяться:
епіназальна (англ.epinasal), яка утворює основу рогу носа;
надвиличні (англ.epijugals) — пара кісток, які часто утворюють невеликі роги вилиці;
надтім'яні (англ.epiparietals) та надлускаті кістки (англ.episquamosals) — невеликі закостеніння, що вкривають краї оборки цератопсів (тім'яна та луската кістки відповідно). Ці окостеніння також називаються надпотиличними (англ.epoccipitals)[29].
епіпофізи (англ.epipophyses) — кісткові виступи шийних хребців, знайдені у динозаврів і деяких викопних базальнихптахів. Ці парні відростки розташовані над постзигапофізами на задній частині невральної дуги хребця. Їхня морфологія дуже різноманітна і варіюється від невеликих, простих, схожих на горбки виступів до великих, складних, крилоподібних наростів. Епіпофізи забезпечували велику площу прикріплення для декількох м'язів шиї. Великі епіпофізи свідчать про сильну мускулатуру шиї. Наявність епіпофізів вважається синапоморфією динозаврів[12].
епіптеригоїд (лат.epipterygoid) — парна ендохондральна кістка в задній частині піднебіння. Невелика, схожа на пластинку кістка, що з'єднується з крилоподібною кісткою та черепною коробкою[12].
задньовушна кістка[30] (англ.opisthotic) — парна кістка задньої частини черепної коробки. У дорослих динозаврів вона завжди зрощена з парними бічними потиличними кістками, утворюючи структуру, відому як бічнопотилично-задньовушна кістка (англ.exoccipital-opisthotic). Нижче парні бічні потиличні зростаються з основною потиличною кісткою[13][12].
зазубрини (англ.serrations) — невеликі виступи, розташовані лініями вздовж ріжучих країв (carinae) зубів. Близьким за значенням є термін «зубчики» (англ.denticle, dentelures). Обидва терміни визначаються по-різному, однак багато авторів використовують їх як синоніми. Інші автори застосовують термін «зазубрини» до менших структур зубів м'ясоїдних тварин, а термін «зубчики» — до грубіших виступів, які можна побачити в багатьох зубах травоїдних тварин. Термінологічний огляд 2015 р. визначає зубчики як складний тип зазубрин, утворених як емаллю, так і дентином, що лежить під нею, тоді як термін «зазубрини» охоплює структури, утворені виключно емаллю. Згідно з цим визначенням, зубні виступи у м'ясоїдних динозаврів, слід називати «зубчиками»[31]. Для опису складної морфології зубів м'ясоїдних використовується детальна термінологія:
*целли (cellae) — проміжки між окремими зубчиками[31].
*діафіз (diaphysis) — точка, де емалеві ковпачки двох сусідніх зубчиків розходяться або знаходяться найближче один до одного[32][33][31].
*ампула (ampulla) — колбоподібна порожнина під діафізом[31].
*корінь (radix) — серцевина зубчика під оперкулумом[31].
*міжзубчикова борозна (interdenticular sulcus чи blood groove) — борозенка, яка проходить від проміжків між сусідніми зубчиками на невеликій відстані до поверхні з обох боків перпендикулярно до ріжучого краю зуба[31].
*cauda — випуклість, що виникає від основи зуба і йде перпендикулярно до ріжучого краю на поверхню зуба між двома міжзубчиковими борознами[31].
зап'ясток Кістки зап'ястя (англ.carpals) утворюють зап'ясток, який з'єднує передпліччя (променеву та ліктьову кістки) з п'ястковими кістками кисті[14]. Відповідною частиною стопи є заплесно (лат.tarsus). Базальні рептилії мають три ряди кісток зап'ястя. У динозаврів зап'ясток часто не повністю закостенілий, а кількість та характер складових зап'ястка переважно залишаються незрозумілими. Особливо добре окостенілий зап'ясток базального птахотазовогогетеродонтозавра складався з дев'яти елементів. Проксимальний ряд складається з os radiale (під променевою кісткою), os ulnare (під ліктьовою кісткою) і горохоподібної кістки (лат.os pisiforme) — невеликого елемента під ліктьовою кісткою та латеральніше за os ulnare. Дистальний ряд складався з п'яти елементів, розташованих над п'ятьма п'ястковими кістками, які позначаються як дистальні п'ясткові кістки 1—5. Середній ряд представлений єдиним елементом — центральною кісткою (лат.centrale)[34][35].
заскроневе вікно (англ.posttemporal fenestra) — парний отвір, розташований під заднім краєм тім'яної частини склепіння черепа і над тім'яним відростком черепної коробки. Можливо, воно було виходом потиличної гілки очної артерії[36].
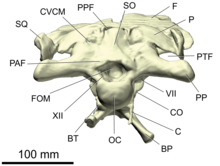
Черепна коробка завроподаSpinophorosaurus у проєкції ззаду, що демонструє затім'яний отвір (PPF) і заскроневе вікно (PTF)затім'яний отвір (англ.postparietal foramen, postparietal fontanelle або postparietal fenestra) — непарний отвір, розташований на середній лінії черепа між тім'яною та верхньою потиличною ділянками, який зустрічається у деяких завроподоморфів. Як і пінеальний отвір, він, можливо, оголював тім'яне око (англ.parietal eye), а отже, слугував для фоторецепції[37][38][39].
зубна батарея Тип зубного ряду у деяких травоїдних динозаврів, де окремі зуби щільно прилягають один до одного, утворюючи безперервну шліфувальну поверхню. У гадрозавраедмонтозавра зубна батарея (англ.dental battery) кожної половини верхньої та нижньої щелеп містила понад 60 рядів зубів. Кожен ряд складався з трьох функціональних зубів, розташованих один на одному, і до п'яти запасних зубів під цим рядом, які прорізалися з щелеп, коли функціональні зуби зношувалися та випадали. Це найскладніший тип зубного ряду у динозаврів — він еволюціонував незалежно у гадрозаврів, цератопсів і деяких завроподів та відрізнявся за формою і функціями в цих окремих кладах[12].
зубні альвеоли, зубні лунки (лат.alveoli) — заглиблення в щелепах, що містять корені зубів. В альвеолах динозаврів зуби періодично замінювалися новими, що росли під ними. Скидання коронки старого зуба відбувалося після резорбції кореня. Стан глибоких альвеол, відомий як текодонтія, присутній у всіх архозаврів, включно з динозаврами. З іншого боку, у більшості лускатих спостерігається плевродонтія, коли зуби зростаються з внутрішніми сторонами кісток щелепи, або акродонтія, за якої зуби зростаються з верхньою частиною кісток щелепи без лунок. Історично стан текодонтії використовувався для визначення класу Thecodontia, який зараз вважається парафілетичним і, отже, застарілим[40].
зчленівна кістка (лат.os articulare) — крайня задня кістка нижньої щелепи. На дорсальній поверхні містить ґленоїд (лат.glenoid) — западину, в яку входить нижній кінець квадратної кістки черепа, утворюючи щелепний суглоб. Це єдина ендохондральна кістка нижньої щелепи, що формується безпосередньо із задньої частини меккелівого хряща. У ссавців зчленована кістка мігрувала у череп, утворивши молоточоксереднього вуха[41].
зчленування (лат.articulatio, англ.articulation) — рухоме з'єднання кісток скелета. Цей термін також використовується для опису збереження зразків — у зчленованому зразку окремі кістки з'єднані разом у своєму оригінальному анатомічному положенні. І навпаки, у розчленованому скелеті кістки виведені зі свого первинного анатомічного положення. Зразок, знайдений з розчленованими кістками, але в безпосередній близькості одна від одної, називається зв'язаним скелетом. Перехід від повністю зчленованого скелета до скупчення ізольованих і незчленованих кісток відбувається поступово[42].
квадратна кістка (англ.quadrate) — велика, парна та ендохондральна кістка в задній частині черепа. Стовпчаста структура, її нижній кінець зчленовується з нижньою щелепою, утворюючи щелепний суглоб, тоді як верхній кінець заходить під лускату кістку. Квадратна кістка є частиною піднебіння[12].
квадратно-вилична кістка (англ.quadratojugal) — кістка в скроневій частині черепа, яку видно збоку, що утворює нижній задній кут черепа[12].
кисть Кисть або рука, складається з кісток зап'ястка, п'ястка та кистьових фаланг пальців.
Схема передньої кінцівки алозавра з основними сегментами кінцівкикінцівки. Парні придатки тіла у тварин, відокремлені від тулуба або голови, разом з поясами, складають частину додаткового скелета. Передні та задні кінцівки дуже схожі за будовою і розділені на три основні частини[15]. Верхня частина, стилоподій, складається з плечової кістки (передня кінцівка) або стегнової кістки (задня кінцівка), тоді як нижня частина, зейгоподій, складається з ліктьової та променевої кісток (передня кінцівка) або великогомілкової та малогомілкової кісток (задня кінцівка). Дистальна частина — автоподій, що складається з кистей або стоп. Сам автоподій поділяється на три сегменти. Зап'ясток (передня кінцівка) та заплесно (задня кінцівка) утворюють базиподій. П'ясток (передня кінцівка) та плесно (задня кінцівка) — метаподій, а фаланги пальців — акроподій[43]. Також, нова термінологія була запропонована Гюнтером Вагнером та Чі-Хуа Чіу у 2001 році. Згідно з нею, автоподій поділяється лише на два відділи, мезоподій, що складається з вузлових елементів (зап'ясток або заплесно), і акроподій, який за цим визначенням включає всі автоподієві довгі кістки, в тому числі п'ясткові кістки (або плеснові кістки) і фаланги[44].
кістки Кістки, окрім зубів, є найпоширенішими скам'янілостями динозаврів. Кістки складаються з гідроксилапатиту, мають сильну мінералізацію і тому міцні. М'які частини, навпаки, можуть зберегтися лише у вигляді відбитків й лише у виняткових випадках. Кісткові частини включають череп, осьовий (хребет і ребра) і аппендикулярний скелет (плечовий, тазовий пояси та кінцівки)[45]. Більшість кісток є парними, з дзеркально оберненим аналогом на іншій стороні тіла, або непарними, в цьому випадку вони зазвичай лежать на середній лінії тіла і розділені останньою на дві симетричні половини[15]. Крім того, кістки охоплюють ряд елементів, що утворюються на шкірі, таких як черевні ребра (лат.gastralia), кісткові гребені (англ.bony scutes) та шипи[45]. Палеонтологи найчастіше вивчають морфологію кісток, але також їхню гістологію (внутрішню мікроструктуру аж до клітинного рівня) та хімічний склад, що дає важливе розуміння біології динозаврів[46]. Існує два основних типи кісток: дермальні кістки[47] (англ.dermal bones) формуються безпосередньо в дермі (шкірі), та виростають з тонких пластинок[47] та ендохондральні кістки, що утворюється з хрящової тканини, яка осифікується (перетворюється на кістку)[15].
ключиці (англ.clavicula) — пара схожих на розпірки, вигнутих кісток, розташованих над коракоїдом у плечі. Ці покривні кістки[47] (англ.dermal bones) прикріплені до акроміоналопатки, і їх найкраще видно на скелеті спереду. Ключиці нечасто знаходять у скелетах динозаврів, що, ймовірно, пов'язано з їхнім низьким потенціалом збереження[49][12].
{{якір|кульшова западина}}{{term|[[кульшова западина]]}} {{defn|кульшова западина, вертлюжна западина (лат.acetabulum) — отвір у тазу, утворений клубовою кісткою, лобком і сідничною кісткою, що видно в бічній і медіальній проєкціях. У ній розміщується головка стегнової кістки, утворюючи тазостегновий суглоб. Більшість тетраподів мають закриту вертлюжну западину, в якій западина повністю заповнена кісткою, утворюючи заглиблення. Динозаври унікальні тим, що демонструють перфоровану або відкриту вертлюжну западину, де вся западина являє собою отвір без заповнення кісткою[15].}}
{{Glossary end}}
кутова кістка Велика кістка нижньої щелепи. У бічному ракурсі кутова кістка (англ.angular) охоплює більшу частину задньої частини щелепи, розташовуючись позаду зубної та нижче надкутової кісток. На медіальному зрізі нижньої щелепи вона розташована нижче передсуглобової кістки. Вона формує дно привідного отвору й підтримує задню частину меккелівого хряща[41].
ліктьова кістка (англ.ulna) — більша і задня з двох кісток передпліччя[15]. На верхньому кінці ліктьової кістки часто видно великий відросток, що тягнеться за ліктьовий суглоб, — ліктьовий відросток (англ.olecranon process), до якого прикріплювалися триголові м'язи (м'язи-розгиначі передпліччя). Іноді значне подовження ліктьвого відростка надавало цим м'язам збільшеного важеля сили[12][2].
лобкова кістка *pubis. Одна з трьох кісток, що складають таз, розташована вентрально від клубової кістки та спереду від сідничної кістки. У ящеротазових лобкове тіло базально спрямоване вперед (пропубічний таз (англ.propubic)). У птахотазових лобкове тіло було спрямоване назад (опістопубічний таз (англ.opisthopubic)) й натомість була присутня добре розвинена передня гілка або препубіс (англ.prepubis), який у похідних формах міг бути більшим за задньоспрямоване тіло[50]. Лівий і правий лобок зазвичай зростаються по всій довжині, а дистальна частина розширюється, утворюючи лобковий фартух (англ.pubic apron)[51]. У деяких неотероподів дистальний відділ шва не є безперервним, а утворює фенестру — лобковий отвір, який видно у черевній проєкції[51]. У проксимальній частині лобка є затульний отвір (англ.obturator foramen), розташований близько до кульшової западини й видимий у бічній проєкції[51]. Затульний отвір забезпечує проходження затульного нерва. У тероподів затульний отвір не повністю оточений кісткою, оскільки бракує вентральної межі; такий стан називається затульною впадиною[51].
лопатка *scapula Ендохондральна кістка і найбільший елемент грудного поясу. У дорослих особин лопатка зазвичай зростається з дзьобоподібною кісткою, утворюючи скапулокоракоїд (англ.scapulocoracoid)[26].
мозочок (лат.cerebellum, букв. «маленький мозок») — дорсальна частина мозку між стовбуром головного мозку і головним мозком, що служить для контролю рівноваги, положення тіла та рухів. Цю частину мозку зазвичай не можна побачити у скам'янілостях динозаврів, оскільки вона рідко зустрічається на ендокранах. Виняток становить можливе збереження мозочкових листків у зразка конхораптора[28][52][14].
надп'яткова кістка. Велика кістка заплесна. Вона розташована безпосередньо під великогомілковою кісткою і медіально від п'яткової кістки, яка знаходиться під малогомілковою кісткою. Надп'яткова кістка (лат.astragalus) не обертається відносно п'яткової або великогомілкової кісток; натомість у двоногих вона часто зливається з обома елементами. Тоді такий елемент скелета називається тибіотарсус (лат.tibiotarsus)[53].
надскроневий отвір надскроневий або верхній скроневий отвір (англ.supratemporal fenestra)
невральна дуга[48][54] (англ.neural arch) — дугоподібний верхній компонент хребця, що лежить над хребетним каналом і тілом хребця. У дорослих динозаврів тіло хребця і невральна дуга зазвичай зростаються разом, закриваючи невроцентральний шов (англ.neurocentral suture), присутній у молодих особин. Невральна дуга може містити різноманітні кісткові відростки, такі як спинні гребені, епіпофізи, поперечні відростки (які з'єднуються з ребрами) і суглобові відростки (що зчленовуються з сусідніми хребцями). Динозаври мають складні невральні дуги, часто оздоблені системою пластинок, ямок та/або плевроцелей, що формують повітряні мішки, які розташовуються вздовж хребців.
носова кістка Парна носова кістка (англ.nasal) є передньою кісткою склепіння черепа і найбільшою кісткою верхньої поверхні морди. Вона розташована між передщелепною спереду і лобовою кісткою ззаду[12][15].
основна клиноподібна кістка (лат.basisphenoideum)[30][47] — кістка черепа, що утворює передню частину дна черепної коробки. Невидима ззовні в зчленованому черепі, вона зрощена з основною потиличною кісткою ззаду і з парасфеноїдом спереду. Пара характерних відростків, основних крилоподібних відростків (лат.processi basipterygoidei), виходять з її нижньої сторони та зчленуються з крилоподібною кісткою піднебіння (лат.pterygoideum)[13].
основна потилична кістка (лат.basioccipitale)[30][47] — непарна кістка черепа, одна з чотирьох кісток, що утворюють потиличну частину черепної коробки. Її видно ззаду на зчленованому черепі. Вона формує більшу частину потиличного виростка (лат.condylus occipitalis), округлого відростка, що з'єднує череп з атлантом, першим хребцем шиї. Зверху зростається з бічними потиличними кістками (англ.exoccipitals) по обидва боки великого отвору черепа[13].
осьовий хребець (лат.axis) — другий хребець хребта, за атласом. Він більший за останній, його центр злитий з невральною дугою (англ.neural arch), хребцевий відросток (англ.neural spine) добре розвинений. Невеликий інтерцентр (лат.intercentrum), розташований перед тілом хребця (лат.centrum), є окремим у тероподів, але зливався з тілом атласу в птахотазових динозаврів[8].
Черепна діаграма базальногозавроподоморфамассоспонділа, що демонструє основні черепні отвори (вікна, ями), разом з зовнішньою ніздрею, передочним отвором, очницею, підскроневим отвором (тут: латеральний скроневий отвір), надскроневим отвором та нижньо-щелепним отвором
передочний отвір (лат.antorbital fenestra) — один з п'яти основних отворів черепа динозаврів, розташований між очницею і ніздрею. Присутній у більшості ранніх архозавроподібних (включаючи архозаврів). Примітивно присутній у динозаврів — він, як правило, великий у ящеротазових, але зменшений або повністю закритий в птахотазових. Передочне вікно лежить у більшому заглибленні — передочній ямці (лат.antorbital fossa). Інколи передочна ямка може містити додаткові менші отвори — верхньощелепний (лат.maxillary fenestra) і передщелепний отвори (лат.promaxillary fenestra)[15].
плевроцелі (англ.pleurocoel) — (Мякушко. Передбачалося, що тіло хребця утворене основами всіх чотирьох елементів дуг (відомих, наприклад, у акули). Ці основи дуг отримали особливі назви — гіпоцентр (основа нижньої дуги) і плевроцентр (його вважали за основу верхньої вставної пластинки))
повітряні мішки У сучасних птахів легеневі повітряні мішки або повітроносні мішки (англ.air sacs) — це тонкостінні, напівпрозорі мішечки, наповнені повітрям, з'єднані з легенями. Разом з легенями повітряні мішки утворюють високоефективну дихальну систему, яка у птахів здатна видобувати до 160 % більше кисню, ніж це можливо у ссавців. Хоча в повітряних мішках відбувається лише обмежений газообмін, вони забезпечують вентиляцію легень. Повітряні мішки, розташовані як ззаду, так і спереду, забезпечують постійний потік повітря через легені.
Менші й сліпі мішечки, наповнені повітрям, що відходять від повітряних мішків і легень, відомі як пневматичні дивертикули (англ.pneumatic diverticula). Вони можуть бути численними й присутніми в більшій частині тіла. Деякі пневматичні дивертикули розсмоктуються і проникають у кістки в міру росту тварини, створюючи заглиблення та порожнини в кістках — процес, відомий як посткраніальна скелетна пневматизація. Хоча повітряні мішки не фосилізуються, на їхню наявність, принаймні у завроподів, вказують характерні сліди пневматизації в кістках[55].
поза смерті (англ.death pose) або опістотонічна поза (англ.opisthotonic posture) — поширена поза, що спостерігається в скам'янілостях динозаврів та птахів, з закинутою назад головою і широко відкритим ротом[56]. Існують різні можливі причини, серед яких опістотонус (передсмертні муки)[56] та розрив еластичної зв'язки ligamentum elasticum під час розкладання тіла[57].
потилиця (лат.occiput) — ділянка на задній стороні черепа[14]. Вона складається з чотирьох кісток, що оточують великий потиличний отвір (лат.foramen magnum) — отвір до спинного мозку: верхня потилична кістка (англ.supraoccipital), бічні потиличні кістки (англ.exoccipitals) та основна потилична (англ.basioccipital) знизу[30]. Безпосередньо під великим потиличним отвором розташований непарний потиличний виріст (англ.occipital condyles), який з'єднується з атлантом, першим хребцем шиї. Потиличний виріст утворений основною потиличною кісткою, й меншою мірою — парними боковими кістками з верхнього краю[13][12].
потилична зв'язка (англ.nuchal ligament, від nuchal, «потилиця, задня частина шиї») —
провідний отвір У рептилій і динозаврів провідний або меккелів отвір (англ.Meckelian orifice, adductor fossa) — великий отвір у нижній щелепі, розташований між зубоносною ділянкою і щелепним зчленуванням. Вона відкривається дорсально. Латерально обмежена надкутовою кісткою, а медіально — передсуглобовою. Оскільки остання зазвичай значно нижча за першу, отвір видно в медіальній проєкції. Дно отвору утворене кутовою кісткою і вміщує задню частину меккелівого хряща. Нижньощелепний отвір та його навколишні краї слугують точкою прикріплення основних привідних м'язів, що закривають щелепу; він також дає змогу головному нерву, артерії та вені щелепи проникати всередину нижньої щелепи[41].
п'яткова кістка (лат.calcaneum) — велика кістка передплесна (в задній частині стопи) і разом з таранною кісткою (лат.talus, astragalus) утворює верхній рівень передплеснових кісток. Вона розташована латерально від таранної і дистально від малогомілкової кістки[53].
стегнова кістка. Проксимальний елемент задньої кінцівки. Головка стегнової кістки спрямована медіально й утворює кут у 90 °, зчленовуючись із кульшовою западиною й утворюючи тазостегновий суглоб[12]. Верхня частина стегнової кістки має низку вертилюгів — відростків для прикріплення м'язів (англ.trochanters):
*менший вертлюг (або передній вертилюг), розташований на передній поверхні безпосередньо під голівкою. Таке розташування є унікальною особливістю для динозаврів і найближчих до них груп. До малого вертлюга кріпиться лобково-сіднично-стегновий м'яз, який тягне задню кінцівку вперед і всередину, і тому має вирішальне значення для пересування. Багато ранніх динозаврів додатково мали кістковий гребінь або м'язовий рубець, відомий як вертилюжна полиця (англ.trochanteric shelf), що тягнувся від основи переднього вертлюга до задньолатерального краю стегнової кістки. Малий вертлюг динозаврів не є гомологічним малому вертлюгу ссавців[12][2].
*більший вертлюг (або дорсолатеральний вертилюг) — гребінь або опуклість на бічній поверхні, яка спрямована протилежно і продовжує головку стегнової кістки[58]. Він забезпечував фіксацію як лобково-стегнового, так і сіднично-вертилюгового м'язів[2].
*четвертий вертлюг — великий фланець на задній поверхні стегнової кістки, спрямований назад. У динозаврів четвертий вертлюг був чітко асиметричний, з проксимальною частиною більш розширеною, ніж дистальна — це їхня синапоморфія. Четвертий вертлюг забезпечував прикріплення довгого хвостово-стегнового м'яза[59], найважливішого рухового м'яза, який при скороченні тягне стегнову кістку назад, забезпечуючи рухову активність[12].
стопа чи нога (англ.pes) — складається з передплесна, плесна та педальних фаланг[15][48]. Перший палець стопи — великий палець, який у більшості двоногих динозаврів був коротким і не контактував з землею, а у птахів — збільшений і обернений (спрямований назад, а не вперед)[1][60].
суглобові відростки (англ.zygapophyses або articular process) — відростки хребетної дуги, що з'єднують сусідні хребці. Кожен хребець має дві пари зигапофізів, які зчленовуються з зигапофізами наступного та попереднього хребців. Спереду розташовані презигапофізи чи краніальні зигапофізи (prezygapophyses, cranial zygapophyses), витягнуті вперед і направлені суглобовою поверхнею догори. На задній частині хребця знаходяться постзигапофізи чи каудальні зигапофізи (postzygapophyses, caudal zygapophyses), які відходять назад суглобовою поверхнею донизу[15].
Класифікація морфологій тіл хребців динозаврів на основі їхніх суглобових зчленуваньтіло хребця (лат.centrum) — елемент у формі котушки або циліндра, який разом з невральною дугою (англ.neural arch) утворює хребець[15]. У молодих особин тіло і дуги хребця є окремими елементами, і їхня зрощеність є важливим критерієм для визначення віку тварини. Передня і задня поверхні тіла утворюють зчленування з центрами попереднього і наступного хребців. Тіла хребців можна класифікувати на основі морфології їхніх суглобових поверхонь:
процельні (лат.procoelous) — увігнуті спереду та випуклі ззаду.
опістоцельні (лат.opisthocoelous) — опуклі спереду та ввігнуті ззаду.
платицельні (лат.platycoelous) — злегка ввігнуті з обох сторін.
амфіцельні (лат.amphicoelous), двояковвігнуті або двобічновигнуті — сильно увігнуті спереду і ззаду.
амфіплатичні (лат.amphiplaty) або ацельні (лат.acoelous) — плоскі з обох кінців, характерні для переважної більшості ссавців[61].
гетероцельні (лат.heterocoelous) або сідлоподібні — мають S-подібну форму з обох кінців[47].
Амфіцельність є первісним станом чотириногих. У риб кінці тіл глибоко увігнуті та з'єднані невеликим отвором — проходом для нотохорди. У рептилій цей тип тіл зустрічається в ембріонів і у дорослих форм деяких видів; у більшості видів, включаючи динозаврів, центри більш окостенілі, а нотохордальний отвір закритий, що забезпечує стійкість до сил стиснення. Гетероцельні хребці забезпечують гнучкість, запобігаючи обертанню. Процельні та опістоцельні тіла хребців утворюють увігнуто-опуклі суглоби, в яких опуклий кінець, виросток (англ.condyle), входить в увігнутий кінець — западину (англ.cotyle). Така конфігурація забезпечує більшу стабільність без обмеження мобільності. У довгих шиях і хвостах ця стабілізація найкраще працює, коли опукла частина спрямована від тіла. Тому у завроподів хребці перед крижем, як правило, були опістоцельними, а хребці хвоста — процельними. Оскільки хребетний стовп може містити хребці різної морфології, утворюються перехідні хребці з двома кінцями різної форми[8][62][63][64].
Скелетна реконструкція пахіцефалозавраHomalocephale, що демонструє його хвостовий кошикхвостовий кошик. сітчаста кістяна структура у хвостах пахіцефалозаврів, що оточує хребетний стовп (але не з'єднується з ним). Хвостовий кошик (англ.caudal basket або myorhabdoi) утворюється в міосепті (сполучній тканині, що розділяє блоки м'язової тканини, міомери) і являють собою окостенілі сухожилля. Тому хвостовий кошик утворюється на периферії м'язової тканини під дермою (а не всередині дерми, як у випадку з остеодермою). Вони також відрізняються від окостенілих сухожиль, які укріплювали хвіст в інших птахотазових, хоча, можливо, виконували схожі функції. Хвостові кошики в першу чергу представлені у костистих риб. Пахіцефалозаври — єдині відомі тетраподи, у яких ці структури також закостеніли. Зчленований хвостовий кошик був виявлений у гомалоцефалів[65].
хвостові хребці Хребці, з яких складається хвіст[14]. У предків динозаврів було приблизно 50 хвостових хребців (англ.caudal vertebrae), хоча їхня кількість, розмір і форма значно варіювалися в окремих групах. Кількість хвостових хребців зменшувалася впродовж еволюційної лінії — у сучасних птахів хвостові хребці, що залишилися, зрослися в пігостиль. Хоча у ранніх динозаврів хвіст був гнучким, у деяких кладах він став жорстким завдяки окостенілим сухожиллям (як у багатьох птахотазових) або подовженим презигапофізам (як у деяких тероподів, таких як дромеозавриди). Диплодоковізавроподи мали видовжений хвіст, схожий на батіг, тоді як хвости деяких птахотазових були вкриті булавами, шипами та/або пластинами[12].
Поперечний розріз хвоста карнотавра, що демонструє хвостову мускулатуру
хвостово-стегнові м'язи (англ.caudofemoralis muscles) — головні рухові м'язи у всіх довгохвостих динозаврів. Розташовані переважно у хвості, вони тягнуть стегнову кістку задньої кінцівки назад, коли скорочуються (втягування стегнової кістки/розгинання стегна), забезпечуючи таким чином рух. У тиранозаврів хвостово-стегновий м'яз був надзвичайно великий і міг становити 58 % від загальної маси хвоста. У динозаврів прослідковувались дві чітко виражені частини: m. caudofemoralis brevis, що бере свій початок на нижньому краї задньої частини клубової кістки, тоді як m. caudofemoralis longus бере початок на передній частині хвоста, вентрально від поперечних відростків і під поверхневими гіпаксіальними м'язами хвоста. Обидві частини кріпилися до четвертого вертлюга на задній поверхні стегнової кістки. Хвостово-стегнові м'язи редукувалися під час еволюції тероподів, і у сучасних птахів вони здебільшого втрачені. Птахи, на відміну від довгохвостих динозаврів, не покладаються на втягування стегнової кістки для руху вперед, а замість цього відтягують гомілку навколо колінного суглоба[2][66].
хоани (лат.choanae), внутрішні носові отвори (англ.internal nares) — пара отворів у піднебінні ротової порожнини, які нерозривно пов'язані із зовнішніми носовими отворами, утворюючи носовий прохід. Лівий і правий отвори розділені по середній лінії черепа лемішем (лат.vomer). У динозаврів хоани зазвичай дуже великі та витягнуті[12][67].
череп охоплює кістки та зуби голови. Він складається з верхньої частини — власне черепа (англ.cranium) та нижньої частини — нижньої щелепи. Череп складається з мозкової коробки, склепіння черепа, верхньої щелепи, області щік та піднебіння. Черепна коробка майже повністю має ендохондральне походження, тоді як решта черепа складається переважно з дермальних кісток. Нижні щелепи складаються як з дермальних, так і з ендохондральних кісток. Череп має ряд отворів, які є важливими опорними точками в анатомічних описах. До них відносяться очниця, або очна ямка, в якій міститься око, а також зовнішній носовий отвір. Крім того, череп у динозаврів мав додаткові отвори, або фенестри, які були характерні для діапсидних рептилій: передочний отвір (англ.antorbital fenestra), між зовнішньою ніздрею і очницею; підскроневий отвір (англ.infratemporal fenestra) позаду очниці; надскроневий отвір (англ.supratemporal fenestra) на задній частині склепіння черепа[15][12][9].
A: вигляд справа; B: вигляд зліва
A: знизу; B: зверху; C: спереду; D: ззаду
3D-реконструкція черепної коробки массоспонділа з різних ракурсів, і пофарбованими окремими кістками. Абревіатури: bo — осн. потилична; bs — зрощені осн. клиноподібна та парасфеноїд; CN — канал черепного нерва; eo — бічнопотилична-задньовушна; ls — бічна клиноподібна; os — орбітоклиноподібна; pa — тім'яна (вважається частиною склепіння черепа); pr — передньовушна; so — верхня потилична; sta — стремінце.
черепна коробка (лат.neurocranium) — відділ черепа, що вміщує головний мозок. У зчленованому черепі його не видно ззовні, окрім задньої частини, потиличного бугра (лат.occiput). Це складна структура, вона пронизана численними отворами (лат.foramina), що містять кровоносні судини та нерви мозку. Окремі кістки черепної коробки у дорослих зазвичай повністю зростаються, причому розмежування окремих частин часто непомітне. Анатомія черепної коробки динозаврів консервативна, і з цієї причини може використовуватися, щоб робити висновки про спорідненості всередині групи, коли інші особливості скелета зазнали настільки глибоких змін, що їхнє походження більше неможливо простежити. Мозковий череп також дозволяє реконструювати мозок та внутрішнє вухо, роблячи висновки щодо органів чуття та інтелекту динозаврів[13].
черепний кінетизм[54] (англ.cranial kinesis) — здатність частин черепа рухатися одна відносно одної в місцях з'єднання всередині черепа. Хоча черепний кінетизм був запропонований для ряду непташиних таксонів динозаврів, аналіз 2008 року показав, що більшість з цих гіпотез є сумнівними[69]. Форми черепного кінетизму, які, можливо, зустрічалися у динозаврів, охоплюють:
стрептостилія (лат.streptostyly) — рух квадратної щодо лускатої кістки у слуховому суглобі (англ.otic joint). Квадратна кістка здебільшого рухається паралельно серединній лінії черепа, хоча можливі й поперечні рухи[69].
прокінетизм або передній кінетизм (англ.prokinesis) — рух між мозковим черепом і лицьовою частиною черепа у носо-лобному суглобі, або рух між лівою і правою носовою кісткою. Ця форма зустрічається у сучасних птахів[69].
neurokinesis — рух мозкового черепа щодо піднебіння у крилоподібному суглобі (або базальному суглобі)[69].
pleurokinesis (плеврокінетизм) — здатність верхньої щелепи висуватися вперед. Плеврокінетизм був запропонований як новий тип черепного кінетизму птахотазових, що дозволив би використовувати поперечні рухи щелепи при жуванні. Ця складна форма черепного кінетизму вимагає, окрім рухливості верхньої щелепи та лицьової частини черепа, поперечних рухів квадратної кістки відносно лускатої (модифікована форма streptostyly), а також кінетизму базального суглоба[69].
шеврон (англ.chevron) або гемальна дуга (англ.haemal arch) — кістки, прикріплені до нижньої сторони хвостових хребців, що утворюють вентральну поверхню хвоста. Шеврон складається з лівої та правої частин, які зазвичай з'єднані між собою у V-подібній формі, утворюючи великий отвір, видимий спереду або ззаду. Отвори кількох послідовних шевронів утворюють гемальний канал хвоста, який захищає нерви та кровоносні судини[14][12].
шийне напівкільце Поперечно орієнтований, комірцеподібний ряд остеодерм, що захищає верхню частину шиї в анкілозаврів (нижня частина шиї залишається оголеною). Кожна особина мала два шийних півкільця (англ.cervical half-ring), причому переднє було меншим за заднє. Шийні півкільця є синапоморфією анкілозаврів[70].
шийні ребра Кістки, які прикріплюються латеровентрально до шийних хребців. Шийні ребра (англ.cervical ribs) є плезіоморфними для амніот (хоча втрачені у ссавців) і складаються з переднього та заднього відростків. Деякі завроподоморфи, особливо деякі довгошиї завроподи, такі як жирафатитан, мали гіперподовжені шийні ребра із задніми відростками, що перекривали два або три попередні хребці. Гістологічний аналіз кісток показав, що ці видовжені відростки являли собою окостенілі сухожилля, а це означає, що їхні кінці були з'єднані з м'язами. Велика довжина відростків збільшила б відстань між тілом м'яза і хребцем, на який він діє, що дозволило б м'язу розташовуватися ближче до тіла, полегшуючи шию[71][72].Ілюстрація шийних хребців диплодока № 15—2
шийні хребці Шийний відділ хребта[14]. Більшість динозаврів мали від 9 до 10 шийних хребців (англ.cervicals), хоча в деяких групах, включаючи завроподів, їхня кількість була більшою, зокрема внаслідок інтеграції спинних хребців у шию[12].
caputegulum Плоскі кістки, що покривають кістки черепа анкілозаврів. Разом з пірамідальнми рогами вони формують оздоблення черепа. Термін, введений Вільямом Т. Блоузом у 2001 році[73], може використовуватися для позначення елементів, що являють собою як коосифікованіостеодерми, так і черепне оздоблення. Положення капутегул (з лат. — «черепні плитки») на черепі можна визначити за допомогою модифікаторів; наприклад, носові капутегули розташовані на носових кістках[74].
carinae (одн. — carina) — гострі, вузькі емалеві гребені, що утворюють ріжучий край зуба на передньому і задньому краях зубів[31]. Вони зазвичай зустрічаються у хижих динозаврів і часто мають зазубрини[12].
centrocoel Печериста порожнина кісткового мозку всередині центру хребця[58].
dolichoiliac (з грец.dolicho- «довгий» та лат.iliac«клубова кістка») — одна з двох основних конфігурацій кісток тазу у ранніх ящеротазових динозаврів. Тип dolichoiliac характеризується великими передніми та задніми розширеннями клубової кістки, а також подовженим і вузьким лобком і сідничною кісткою. На противагу цьому, тип brachyiliac (з грец.brachy- «короткий» та лат.iliac «клубова кістка») характеризується коротким розширенням клубової кістки та більш міцними лобковими та сідничною кісткою. Тип brachyiliac характерний для базальних завроподоморфних динозаврів тріасу, тоді як тип dolichoiliac характерний для тероподів. Обидва терміни були введені Едвіном Колбертом у 1964 році[76][77].
epijugal або надвилична кістка (з грец.epi- «над» та лат.jugal«вилична кістка») epijugal — унікальне для цератопсів шкірне окостеніння. Воно закривало нижній кінець спрямованого донизу трикутного фланця виличної кістки, характерного для цієї групи. У Arrhinoceratops[en] і Pentaceratops[en] надвилична кістка утворювала чіткий виличний ріг. Ця кістка зустрічалася у ямацератопсів і всіх більш ранніх цератопсів[78][79].
epoccipital або надпотилична кістка (з грец.ep- «над» та лат.occipital«потилична кістка») epoccipital — шкірні окостеніння, що покривали краї оборки цератопсів. Надпотиличні кістки були окремими кістками у молодих особин, але у дорослих зросталися або з лускатою, або з тім'яною кістками, залежно від їхнього розташування. Надпотиличні кістки, що зростаються з лускатою також відомі як надлускаті, а тім'яні — як надтім'яні. Деякі автори виступають проти використання терміну «надпотилична кістка», оскільки ця кістка не пов'язана з потиличною ділянкою черепа, натомість надають перевагу термінам «надлускаті» та «надтім'яні» кістки. Надпотиличні кістки були скоріше декоративними, ніж функціональними, і широко варіювали за формою, наприклад утворюючи великі шипи у центрозаврів[80][78][79].
hypex Вершина кута, утвореного між двома пальцями, або, іншими словами, найближча точка між двома пальцями. Гіпекс можна ідентифікувати лише за наявності м'яких тканин. Цей термін здебільшого застосовують при вивченні відбитків скам'янілостей[81].
Zahnreihe (з нім.«зубний ряд», множ. — Zahnreihen) — ряд зубних одиниць, що утворюють одиницю заміщення. Зубний ряд, як правило, складається з декількох Zahnreihe, що містять собою по два чи більше зубів. Заміна зубів завжди починається з найвіддаленішого заднього зуба і продовжується вперед. Таким чином, уникається одночасна заміна двох сусідніх зубів, що може призвести до утворення проміжків у зубному ряді. Кількість зубів від одного Zahnreihe до іншого є крок-Z. Коли крок-Z дорівнює рівно 2, зуби замінюються по черзі; якщо крок-Z більший за 2, зуби замінюються послідовно[82].
ziphodont Лезоподібні зуби з вигнутими та сплющеними коронками та, як правило, зазубринами. Пристосовані до м'ясоїдної дієти, цей тип зубів базально присутній у динозаврів і зустрічався у більшості непташиних тероподів[75].
↑ абвгдеDilkes, D. W.; Hutchinson, J. R.; Holliday, C. M.; Witmer, L. M. (2012). Reconstructing the musculature of dinosaurs. The Complete Dinosaur (англ.). Bloomington: Indiana University Press. с. 151—190.
↑Leonardi, Giuseppe (1987). Glossary and manual of tetrapod footprint palaeoichnology. Brasília: Publicação do Departemento Nacional da Produção Mineral Brasil. с. 43—51.
↑ абвгRomer, A.S. (1976). The Axial Skeleton. Osteology of the Reptiles(англ.). The University of Chicago Press. с. 218—297. ISBN978-0-226-72491-1.
↑ абвRomer, A.S. (1976). The Skull–General Structure. Osteology of the Reptiles. The University of Chicago Press. с. 55—83. ISBN978-0-226-72491-1.
↑Romer, A.S. (1976). The Skull in the Various Reptile Groups. Osteology of the Reptiles. The University of Chicago Press. с. 85—197. ISBN978-0-226-72491-1.
↑ абвгдежикBrett-Surman, M.K.; Holtz, T.R.; Farlow, J.O., ред. (2012). Glossary. The Complete Dinosaur (вид. друге). Indiana University Press. с. 1075—1081. ISBN978-0-253-00849-7.
↑ абвгдежиклмнпрстHoltz, Thomas R. Jr.; Brett-Surman, Michael K. (1997). Farlow, J. O.; Brett-Surman, Michael K. (ред.). The Osteology of the Dinosaurs. The Complete Dinosaur (англ.). Bloomington: Indiana University Press. с. 78—91.
↑Novas, Fernando E. (2009). The Age of Dinosaurs in South America. Nature (англ.). Bloomington: Indiana University Press. с. 452. ISBN978-0-253-35289-7.
↑Upchurch, P.; Barrett, P.M.; Dodson, P. (2004). Sauropoda. У Weishampel, D.B.; Dodson, P.; Osmolska, H. (ред.). The Dinosauria, Second Edition. Univ of California Press, Berkeley. с. 259—322. ISBN978-0-520-24209-8.
↑Wu, Xiao-Chun; Russell, Anthony P. (1997). Functional Morphology. У Kevin Padian; Philip J. Currie (ред.). Encyclopedia of dinosaurs(PDF). San Diego: Academic Press. с. 258—268. ISBN978-0-12-226810-6.
↑Fastovsky, David E.; Weishampel, David B. (2009). Dinosaurs: a concise natural history(англ.). Cambridge: Cambridge University Press. с. 358. ISBN978-0-521-88996-4.
↑ абVickaryous, M.K.; Maryańska, T.M.; Weishampel, D.B. (2004). Ankylosauria. У David B. Weishampel; Peter Dodson; H. Osmólska (ред.). The Dinosauria (вид. друге). Berkeley: University of California Press. с. 363—392. ISBN978-0-520-24209-8.
↑Holtz JR., T.R.; Molnar, R.E.; Currie, P.J. (2004). Basal Tetanurae. У David B. Weishampel; Peter Dodson; H. Osmólska (ред.). The Dinosauria (вид. 2nd). Berkeley: University of California Press. с. 71—110. ISBN978-0-520-24209-8.
↑Galton, P.M. (2012). Stegosaurs. У Brett-Surman, M.K.; Holtz, T.R.; Farlow, J.O. (ред.). The Complete Dinosaur (вид. 2). Indiana University Press. с. 482—504. ISBN978-0-253-00849-7.
↑Galton, Peter M.; Upchurch, Paul (2004). Prosauropoda. У David B. Weishampel; Peter Dodson; H. Osmólska (ред.). The Dinosauria (вид. друге). Berkeley: University of California Press. с. 232—258. ISBN978-0-520-24209-8.
↑ абвгдБулахов, В. Л.; Новіцький, Р. О.; Гассо, В. Я.; Пахомов, О. Є. (2009). Зоологія хордових(PDF)(укр.). Дніпро: Дніпровський національний університет імені Олеся Гончара. с. 128.
↑Läng, Émilie; Goussard, Florent (2007). Redescription of the wrist and manus of ?Bothriospondylus madagascariensis: new data on carpus morphology in Sauropoda. Geodiversitas. 29 (4): 549—560.
↑Langer, Max C. (2004). Basal Saurischia. У David B. Weishampel; Peter Dodson; H. Osmólska (ред.). The Dinosauria (вид. 2nd). Berkeley: University of California Press. с. 25—46. ISBN978-0-520-24209-8.
↑Schünke, M.; Schulte, E.; Schumacher, U. (2006). Ross, L.M.; Lamperti, E.D. (ред.). Thieme Atlas of Anatomy: General Anatomy and Musculoskeletal System. Thieme. ISBN978-3-13-142071-8.
↑ абвгдежиклМарисова, І. В.; Кузьменко, Л. П.; Власенко, Р. П. (2016). Зоологія хордових(PDF) (навчальний посібник) (укр.). Ніжин: НДУ ім. М. Гоголя. с. 208.
↑ абвгдЦарик, Й. В.; Хамар, І. С.; Дикий, І. В. (2013). Царик, Й. В. (ред.). Зоологія хордових(PDF) (підручник). Біологічні Студії (укр.). Львів: ЛНУ ім. Івана Франка. с. 356. ISBN978-966-613-752-7.
↑ абMadsen Jr., J.H.; Welles, S.P. (2000). Ceratosaurus (Dinosauria, Theropoda) a Revised Osteology(англ.). Utah Geological Survey. с. 1—80. ISBN978-1-55791-380-7.
↑Омельковець, Я.А.; Сологор, К.А.; Білецька, М.Г.; Лихотоп, Р.Й.; Пикалюк, В.С.; Шварц, Л.О. (2003). Порівняльна анатомія хребетних тварин(PDF) (навчальний посібник) (укр.). Луцьк: РВВ «Вежа» Волинського державного університету ім. Лесі Українки. с. 208. ISBN966-600-120-9.
↑Naish, D. (2012). Birds. У Michael K. Brett-Surman; Thomas R. Holtz; James Orville Farlow (ред.). The complete dinosaur. Indiana University Press. с. 379—423.
↑Allaby, Michael (2014). A Dictionary of Zoology(англ.) (вид. четверте). Oxford: Oxford University Press. ISBN978-0-19-968427-4.
↑Kardong, Kenneth V. Skeletal System: The Axial Skeleton. Vertebrates – Comparative Anatomy, Function, Evolution (вид. 6.). Mc Graw Hill. с. 294—324. ISBN978-9-33-922211-6.
↑Carpenter, K.; Tidwell, V. (2005). Reassessment of the Early Cretaceous sauropod Astrodon johnsoni Leidy 1865 (Titanosauriformes). У Carpenter, K.; Tidwell, K. (ред.). Thunder-lizards: the sauropodomorph dinosaurs. Indiana University Press. с. 78—114. ISBN978-0-253-34542-4.
↑William T., Blows (2001). Dermal Armor of the Polacanthine Dinosaurs. У Carpenter, Kenneth (ред.). The Armored Dinosaurs. The Life of the Past (англ.). Bloomington, Ind: Indiana University Press. с. 363—385. ISBN978-0-253-33964-5.
↑ абDodson, P.; Forster, C. A.; Sampson, S. D. (2004), Ceratopsidae, у Weishampel, D. B.; Dodson, P.; Osmólska, H. (ред.), The Dinosauria (вид. 2), Berkeley: University of California Press, с. 494—513, ISBN978-0520254084
↑ абMakovicky, P. (2012). Marginocephalia. У Brett-Surman, M.K.; Holtz, T.R.; Farlow, J.O. (ред.). The Complete Dinosaur (вид. 2). Indiana University Press. с. 526—549. ISBN978-0-253-00849-7.
↑Leonardi, Giuseppe (1987). Glossary and manual of tetrapod footprint palaeoichnology. Brasília: Publicação do Departemento Nacional da Produção Mineral Brasil. с. 43—51.